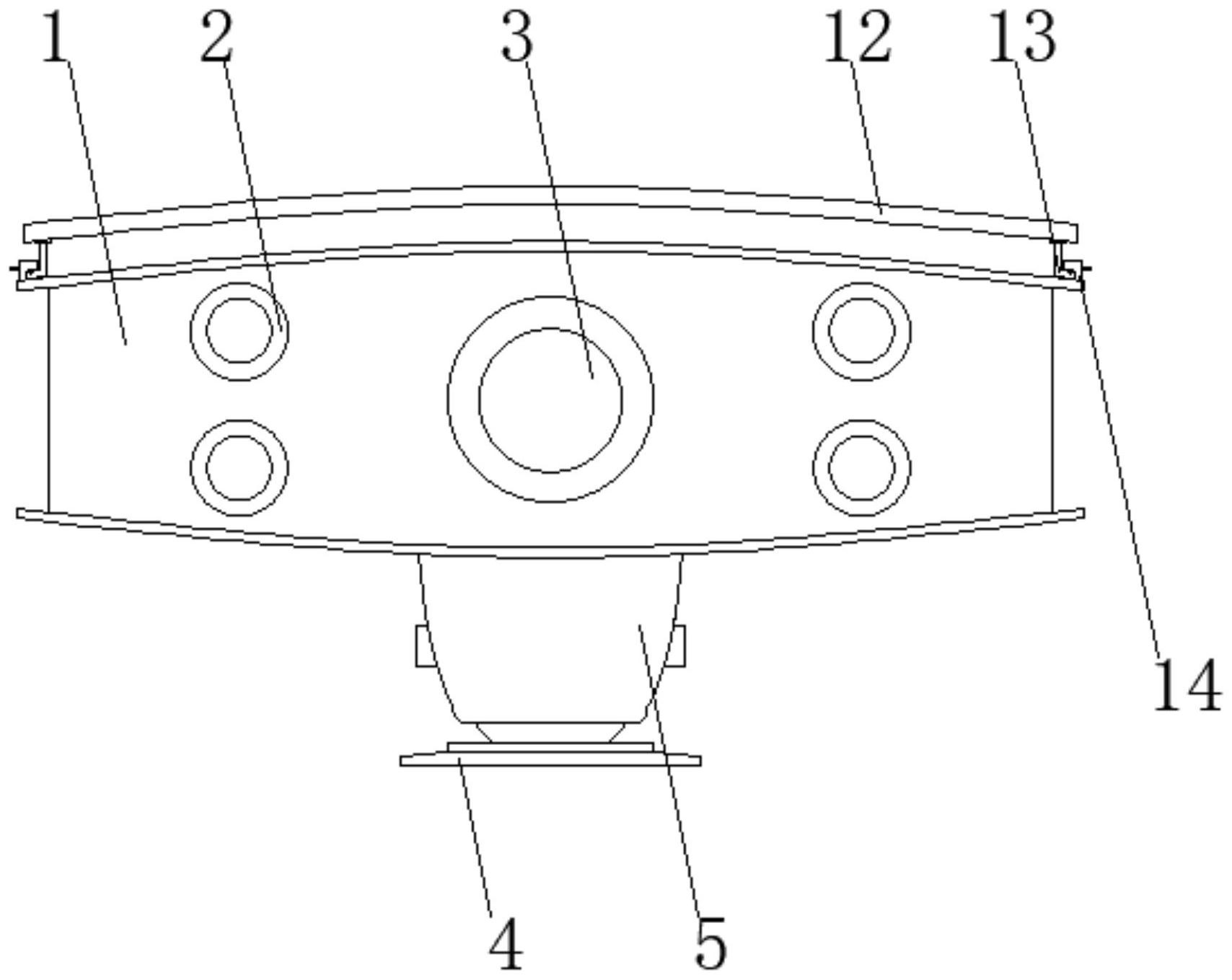
技术摘要:
提供了用于确定细菌对抗生素剂的敏感性或抗性的方法。在一个实施方案中,所述方法包括在抗微生物剂存在或不存在下培养细菌以产生原代培养物,其然后在转化噬菌体存在或不存在下培养。重组噬菌体对细菌是特异性的,并且包含异源标志物(例如,可表达为可检测产物如RNA或 全部
背景技术:
自抗生素青霉素首次实际应用以来,已经开发出许多其他抗菌剂,且抗菌疗法极 大地促进了现代医学的进步和平均寿命的延长。然而,致病细菌已经获得对大多数抗菌剂 的抗性,从而损害了抗菌疗法的整体有效性,同时也带来了新的公共健康问题。特别地,对 β-内酰胺抗菌剂具有抗性的耐甲氧西林金黄色葡萄球菌(MRSA)是高度抗性的病原体。它与 每年近94 ,000例新住院病例直接相关,仅在美国就导致约19 ,000例死亡/年(Voss等, International Journal of Antimicrobial Agents,5:101-106,1995;McGeer等,LPTP Newsletter,190:1-4,1996;CDC MRSA tracking)。部分地由于在畜牧业和医院中抗生素的 使用增加,新的耐多药细菌菌株也以惊人的速度出现。例如,已经报道了用万古霉素治疗 MRSA感染的患者中的万古霉素中介金黄色葡萄球菌(VISA)感染(Hiramatsu等,J Antimicrob Chemother .40(1) ,135-6 ,1997;Perichon等,Antimicrob Agents Chemother.,53(11):4580-7,2009)。实际上,一些菌株已经对几乎所有常用药剂产生抗性。 众所周知的情况是MRSA的Mu50菌株,其对氨基糖苷类、大环内酯类、四环素、氯霉素和林可 酰胺也具有抗性(Hiramatsu等,同上)。还鉴定了对异烟肼和利福平抗性的耐多药结核分枝 杆菌(Dalton等,Lancet,380:1406-17,2012)。 食源性细菌性疾病,特别是由耐药细菌引发的那些疾病,也对人类健康构成重大 威胁。一项分析了包括蔬菜沙拉、生鸡蛋表面、生鸡肉、未巴氏消毒的牛奶和生肉的150种食 物样品的检测大肠杆菌的微生物研究表明,在生鸡肉中检测到最高百分比的耐药性大肠杆 菌分离株(23.3%),其次是蔬菜沙拉(20%)、生肉(13.3%)、生鸡蛋表面(10%)和未巴氏消 毒的牛奶(6.7%)。耐药性大肠杆菌的总体发生率为14.7%(Rasheed等,Rev Inst Med Trop Sao Paulo,56(4):341-346,2014)。该研究进一步强调了耐药性大肠杆菌将抗药性基 因转移到其他物种(例如克雷伯氏菌属)的能力所带来的威胁。 越来越多的科学证据表明细菌如何发展防御系统以针对目前使用的五大类抗菌 药物提供保护。这些药物大致分类为β-内酰胺类、β-内酰胺酶抑制剂、头孢菌素类、喹诺酮 类、氨基糖苷类、四环素类/甘氨酰环素类和多粘菌素类。每种药剂的局限性,特别是当单独 4 CN 111601897 A 说 明 书 2/43 页 使用时,概述如下。 β-内酰胺类是一大类广谱药物,其是革兰氏阴性感染的主要治疗方法。β-内酰胺 类药物的亚类范围从窄谱(青霉素)到广谱(碳青霉烯)。革兰氏阴性细菌已经形成了几种β- 内酰胺抗性的途径。也许令人担忧的机制涉及β-内酰胺酶的进化,β-内酰胺酶是破坏β-内 酰胺抗生素的酶。一些β-内酰胺酶破坏窄谱药物(例如,仅对青霉素是活性的),而较新的β- 内酰胺酶(例如在耐碳青霉烯肠杆菌科或CRE中发现的碳青霉烯酶)能够中和所有β-内酰胺 抗生素。 β-内酰胺酶抑制剂仍具有抗革兰氏阴性细菌的活性,所述革兰氏阴性细菌具有的 β-内酰胺酶具有有限的破坏β-内酰胺抗生素的活性。对超广谱头孢菌素类和碳青霉烯类具 有抗性的细菌通常也对这些药物有抗药性。开发中的新的β-内酰胺酶抑制剂组合药物有可 能克服来自最强效的β-内酰胺酶(例如在CRE中发现的那些)的一些(但不是全部)抗性。 在过去的20年中,超广谱头孢菌素类一直是治疗严重革兰氏阴性菌感染的基础。 耐药性革兰氏阴性感染正在蔓延到社区。耐药性通常使碳青霉烯成为唯一有效的抗菌剂。 氟喹诺酮类药物是广谱抗生素,其通常口服给药,使其便于在住院和门诊患者中 使用。然而,随着在患者群体中使用的增加,耐药菌株迅速进化,使药物无效。增加的使用还 与艰难梭菌的耐药性、超毒力菌株引起的感染增加有关。 氨基糖苷类通常与β-内酰胺药物联合用于治疗由革兰氏阴性细菌引起的感染。尽 管存在越来越多的抗药性问题,这些药物作为对抗严重感染的最后手段仍然是一种重要的 治疗选择。然而,由于担心耐药性及其长期副作用,临床医生很少(如果有的话)单独使用它 们。 四环素不是严重革兰氏阴性感染的一线治疗选择;然而,由于其他药物种类的疗 效有限,它们被认为是治疗严重感染的一种选择。甘氨酰环素类(即替加环素)通常被考虑 用于治疗耐多药革兰氏阴性感染。替加环素是一种不在体内均匀分布的药物,因此根据感 染部位常常与其他药物组合使用。尽管相对不常见,但已报道对替加环素具有抗性的菌株 的发生。 由于毒性问题,多粘菌素类是不受欢迎的较老种类。现在它们经常被用作治疗耐 多药革兰氏阴性菌感染的“最后手段”药剂。由于这些是仿制药,因此在剂量学和疗效方面 的当代数据有限。另外,关于高抗性菌株的检测,存在一些但有限的数据。 鉴于耐药细菌菌株数量的快速增加,迫切需要新的和有效的方法来鉴定和核型分 析临床和非临床细菌分离株,特别是属于ESKAPE组(屎肠球菌、金黄色葡萄球菌、肺炎克雷 伯菌、鲍氏不动杆菌、铜绿假单胞菌和肠杆菌属)的那些。(Boucher等,Clinical Infectious Diseases,48:1-12,2009)。还需要快速准确的病原体鉴定以使医生能够对感 染(包括可能危及生命的那些感染)作出反应和适当地应对。目前,病原体鉴定需要在固体 培养基(基于琼脂的平板)上培养,然后进行诊断分析,该诊断分析通常需要在培养中额外 的复制轮或特定细菌产物的纯化。最好的情况下,微生物鉴定需要多天,在此期间可能需要 额外的生物安全防护等级,这取决于病原体的总体分类。这种生物学的、基于生长的分析的 第二代形式通过使用辐射测量(例如,Becton Dickinson的BACTECTM)或比色/荧光测定(例 如,Becton Dickinson的MGITTM和Biomerieux的 )装置以测量生长细菌所产生的 代谢产物而不是等待细菌群体达到足以被祼眼看到的密度来缩短微生物鉴定和抗性测试 5 CN 111601897 A 说 明 书 3/43 页 的检测时间。然而,这些分析系统经常面临污染问题,因此增加了再处理的需要并导致不必 要的延迟(Tortoli等,J.Clin.Microbiol.,40:607-610,2002)。 最近加速药物抗性细菌的生物学检测的方法集中于使用噬菌体来探测抗微生物 剂对分离株的影响(Schofield等,Bacteriophage,2(2):105-283,2012和WO 08/124119)。 噬菌体用于感染细菌,劫持宿主的细胞生物合成机构进行复制,从而作为鉴定临床标本中 特定细菌菌株存在的工具。可以在噬菌体检测中采用多种方法。一种方法依赖于核酸扩增 的使用(美国专利公开号2014-0256664和WO 12/158502)。在该方法中,通过分析分枝杆菌 噬菌体D29 DNA的实时PCR产物来筛选结核分枝杆菌的药物敏感性。 相关方法依赖于用携带噬菌体的细菌感染次级培养物并分析次级培养物的生长 特性。该方法通常用于鉴定药物抗性结核分枝杆菌。基于这种间接检测方法的示例性商业 试剂盒由Biotec,Inc.(Suffolk,UK)以商标FASTPLAQUE-RESPONSETM销售(Mole等,J Med Microbiol .,56(Pt 10):1334-9 ,2007;Albert等,JAppl Microbiol .,103(4):892-9 , 2007)。该试剂盒还提供了分枝杆菌噬菌体D29,然而,与D29 DNA的直接PCR分析相反,该方 法试图通过使用杀病毒剂来消除不感染细菌的噬菌体而最小化假阳性。在筛选感染的分枝 杆菌后,将噬菌体感染的结核分枝杆菌与快速复制的耻垢分枝杆菌混合,且然后将混合物 接种在琼脂板上。该测定系统基于以下原理:耻垢分枝杆菌被D29有效地交叉感染并在耻垢 分枝杆菌细菌菌苔上形成清晰和可见的斑块,使得每个斑块代表最初被D29感染的结核分 枝杆菌细胞。因此,该测定法定量测量小的结核分枝杆菌池中的D29复制。虽然是准确和快 速的测试,但该测定法太复杂和难以在资源贫乏的环境中使用,因为通过琼脂板上的斑块 形成分析病毒生长必须由经训练的技术人员在实验室中进行。此外,可用于该测定的次级 快速生长细菌的数量是有限的,该测定不能定制或修改以筛选大量目标细菌物种。 类似地,原始荧光素酶报告实验(LRA)的变型,例如使用工程化分枝杆菌噬菌体 TM4,在检测灵敏度方面也受到限制。参见,Piuri等,PLoS One,2009;4(3):e4870,其中荧光 噬菌体(fluorophage)(荧光分枝杆菌噬菌体(fluoromycobacteriophage))在感染后16小 时仅能检测到50%的结核分枝杆菌细胞。而且,因为该测定包括检测在小样品中表达的荧 光或发光标记物,所以该测定对于可以分析的样品类型是有限的。 总之,目前鉴定药物抗性细菌的方法不能满足当今对多种多样的细菌(包括其混 合物)的表型分析的高效和有效手段的需要,例如,基于它们所具有的抗性类型。因此迫切 需要可用于筛选特定细菌菌株对抗菌剂的易感性的分析系统。这种分析技术可以有效地与 许多人和兽医学疾病(如霍乱、脑膜炎、肺炎等)的诊断、治疗和控制相结合。这些系统和分 析也可用于筛选可用于补充工业上有用的微生物(例如大肠杆菌、富养罗尔斯通氏菌、肉葡 萄球菌等)的生长的益菌剂(probiotics)。
技术实现要素:
因此,本发明的目的是提供比目前可用技术提供的更低成本、更高效、更特异性、 更快速、更易得且更高适应的用于选择性微生物(例如细菌)检测的方法和设备。因此,提供 了一种用于确定细菌对抗生素的抗性和用于微生物物种鉴定的方法。该方法利用了噬菌体 对其相应宿主细菌的内在特异性。在一个实施方案中,提供了用于鉴定引起感染的细菌物 种并同时确定鉴定的细菌对抗微生物剂或抗生素剂的易感性的方法。 6 CN 111601897 A 说 明 书 4/43 页 根据前述内容,实施方案提供了重组噬菌体、用于构建和产生此类重组噬菌体的 方法以及使用此类重组噬菌体检测目标细菌和/或确定目标细菌对其具有抗性的药物或抗 生素的方法。所述组合物和方法还可以适应于筛选新的益菌剂,其可用于以实验室水平或 工业规模生物合成酶、激素、抗体、核酸、糖和其他生物分子。 根据一个实施方案,能够检测特定类型细菌的产品、试剂盒和方法,例如,通过探 测靶向的活细菌中特定分子(例如标志物,如蛋白质)的存在。一旦鉴定出药物抗性菌株,该 方法可以例如与其他技术相结合用于鉴定药物抗性机制的分子基础,例如基因突变、基因 复制、转化、抗生素降解等。目前利用包含可通过免疫测定检测的异源肽/蛋白质标志物的 基因的重组噬菌体实现了上述目的。 在一个实施方案中,提供了一种用于鉴定样品中细菌物种的方法。该方法包括将 样品或样品的等分试样与经转化以表达异源蛋白质标志物的噬菌体一起培养或孵育以形 成转化的培养物,并测定转化的培养物中异源蛋白质标志物的存在或不存在。标志物的存 在表明该细菌物种的存在。在一个实施方案中,使用侧流免疫分析进行测定。 在一个实施方案中,噬菌体选自由裂解性噬菌体、溶原性噬菌体和丝状噬菌体组 成的组。 在另一实施方案中,裂解性噬菌体选自由T4、T7、T3和MS2组成的组。 在另一实施方案中,溶原性噬菌体是λ噬菌体。 在另一实施方案中,丝状噬菌体选自由fl、fd和M13组成的组。 在另一实施方案中,标志物在细菌中可表达成核酸或蛋白质。 在另一实施方案中,标志物可表达成选自由抗原、酶、抗体或其片段和适体,或其 组合组成的组的多肽。 在另一实施方案中,蛋白质标志物包含可检测的标记。 在另一实施方案中,蛋白质标志物或标志物上的可检测的标记用选自荧光分析、 化学发光分析、酶分析、凝胶电泳、免疫分析和配体结合分析的分析法进行检测。 在另一实施方案中,蛋白质标志物上的可检测的标记用侧流免疫分析进行检测。 在另一实施方案中,孵育还包括在抗微生物剂存在下的孵育,其中异源蛋白质标 志物的表达指示对抗微生物剂的细菌抗性。 在另一方面,提供了用于同时鉴定样品中的细菌物种和确定其对抗微生物剂的易 感性的方法。该方法包括(a)在具有抗微生物剂的情况下培养样品或等分的样品以产生原 代培养物;(b)在具有对细菌物种特异性且经工程化以表达异源标志物的转化噬菌体的情 况下培养原代培养物;和(c)检测标志物的存在或不存在,其中标志物的存在指示所述样品 中所述细菌物种的存在和其对所述抗微生物剂的抗性。 在另一方面,提供了一种用于同时鉴定样品中的细菌物种和确定其对抗微生物剂 的易感性的方法。该方法包括(a)在具有或不具有抗微生物剂的情况下培养等分的样品以 产生一组原代培养物;(b)在具有或不具有对细菌物种特异性且经工程化以表达异源标志 物的转化噬菌体的情况下培养所述组的原代培养物的部分,从而产生多个转化的次级培养 物,其中第一转化次级培养物包括在具有抗微生物剂的情况下培养的转化细菌和第二转化 次级培养物包括不是在具有抗微生物剂的情况下培养的转化细菌;和(c)检测异源标志物 的存在或不存在,其中所述标志物的存在指示样品中细菌物种的存在和其对抗微生物剂的 7 CN 111601897 A 说 明 书 5/43 页 抗性。 在再另一方面,提供了用于确定细菌对测试抗微生物剂的易感性的方法。该方法 包括(a)在抗微生物剂存在和不存在的情况下培养细菌以产生原代培养物;(b)在对细菌特 异性且包含标志物的转化噬菌体存在和不存在的情况下培养原代培养物,从而产生多个转 化的次级培养物,其中第一转化次级培养物包括已经用测试抗微生物剂处理的转化细菌和 第二转化次级培养物包括未用所述抗微生物剂处理的转化细菌;和(c)检测所述第一和第 二转化次级培养物各自的所述标志物的水平或活性,从而确定所述细菌对抗微生物剂的易 感性。 在一个实施方案中,提供了用于确定细菌对测试抗微生物剂的易感性的方法,其 中该方法包括(a)在抗微生物剂存在或不存在的情况下培养细菌以产生多个原代培养物; (b)在对细菌特异性且包含标志物的转化噬菌体存在和不存在的情况下培养(a)的原代培 养物,从而产生多个转化的次级培养物,其中第一转化次级培养物包括已经用所述测试抗 微生物剂处理的转化细菌和第二转化次级培养物包括未用所述抗微生物剂处理的转化细 菌;和(c)检测第一和第二转化次级培养物各自的标志物的水平或活性,从而确定所述细菌 对抗微生物剂的易感性。在这种情况下,步骤(a)、(b)和(c)可以顺序地或非顺序地进行。在 特定的实施方案中,步骤(a)、(b)和(c)顺序地进行。 在相关的实施方案中,提供了用于确定细菌对测试抗微生物剂的易感性的方法。 该方法包括(a)在抗微生物剂存在和不存在的情况下培养细菌以产生多个原代培养物;(b) 在对细菌特异性且包含标志物的转化噬菌体存在或不存在的情况下培养(a)的原代培养 物,从而产生多个次级培养物,其中第一转化次级培养物包括已经用测试抗微生物剂处理 的转化细菌和第二转化次级培养物包括未用抗微生物剂处理的转化细菌;和(c)检测第一 和第二转化次级培养物各自的标志物的水平或活性,其中与第二转化次级培养物(对照)中 标志物的水平或活性相比,第一转化次级培养物中标志物的水平或活性的降低指示细菌对 测试抗微生物剂是易感。 在另一相关实施方案中,提供了用于确定细菌对测试抗微生物剂的易感性的方 法。该方法包括(a)在抗微生物剂存在和/或不存在的情况下培养细菌以产生多个原代培养 物;(b)在对细菌特异性且包含标志物的转化噬菌体存在或不存在的情况下培养(a)的原代 培养物,从而产生多个转化的次级培养物,其中第一转化次级培养物包括已经用测试抗微 生物剂处理的转化细菌和第二转化次级培养物包括未用抗微生物剂处理的转化细菌;和 (c)检测第一和第二转化次级培养物各自的标志物的水平或活性,其中第一转化次级培养 物中标志物的水平或活性与第二转化次级培养物(对照)中标志物的水平或活性相比的一 致(例如,无变化)或提高指示所述细菌对测试抗微生物剂不是易感的或者对测试抗微生物 剂是抗性的。 在另一实施方案中,提供了用于确定测试剂对细菌的益生效果的方法。该方法包 括(a)在所述测试剂存在和/或不存在的情况下培养细菌以产生多个原代培养物;(b)在对 细菌特异性且包含标志物的转化噬菌体存在或不存在的情况下培养(a)的原代培养物,从 而产生多个次级培养物,其中第一转化次级培养物包括已经用测试剂处理的转化细菌和第 二转化次级培养物包括未用测试剂处理的转化细菌;和(c)检测第一和第二转化次级培养 物各自的标志物的水平或活性,其中第一转化次级培养物中标志物的水平或活性与第二转 8 CN 111601897 A 说 明 书 6/43 页 化次级培养物(对照)中标志物的水平或活性相比的提高指示测试剂具有益生效果。 在另一实施方案中,提供了用于确定格兰氏阳性或格兰氏阴性细菌对测试抗微生 物剂的易感性的方法。该方法包括(a)在抗微生物剂存在和/或不存在的情况下培养格兰氏 阳性或格兰氏阴性细菌以产生多个原代培养物;(b)在对格兰氏阳性或格兰氏阴性细菌特 异性且包含标志物的转化噬菌体存在或不存在的情况下培养(a)的原代培养物,从而产生 多个次级培养物,其中第一转化次级培养物包括已经用测试抗微生物剂处理的转化的格兰 氏阳性或格兰氏阴性细菌和第二转化次级培养物包括未用抗微生物剂处理的转化的格兰 氏阳性或格兰氏阴性细菌;和(c)检测第一和第二转化次级培养物各自的标志物的水平或 活性,从而确定格兰氏阳性或格兰氏阴性细菌对抗微生物剂的易感性。 在一个实施方案中,细菌选自由鲍氏不动杆菌(Acinetobacter baumannii)、炭疽 芽孢杆菌(Bacillus anthracis)、蜡状芽孢杆菌(Bacillus cereus)、百日咳博德特氏菌 (Bordetella pertussis)、伯氏疏螺旋体(Borrelia burgdorferi)、流产布鲁氏菌 (Brucella aborus)、犬布鲁氏菌(Brucella canis)、羊布鲁氏菌(Brucella melitensis)、 猪布鲁氏菌(Brucella suis)、空肠弯曲杆菌(Campylobacter jejuni)、肺炎衣原体 (Chlamydia pneumoniae)、鹦鹉热衣原体(Chlamydia psittaci)、沙眼衣原体(Chlamydia trachomatis)、肉毒杆菌(Clostridium botulinum)、艰难梭菌(Clostridium difficile)、 产气荚膜梭菌(Clostridium perfringens)、破伤风梭菌(Clostridium tetani)、白喉棒状 杆菌(Corynebacterium diphtheriae)、肠杆菌属(Enterobacter sp .)、粪肠球菌 (Enterococcus faecalis)、耐万古霉素粪肠球菌(vancomycin-resistant Enterococcus faecalis)、屎肠球菌(Enterococcs faecium)、大肠杆菌(Escherichia coli)、产肠毒素大 肠杆菌(en te ro to x ig e n i c Es c h e ri c h ia c o l i ,E T E C)、肠致病性大肠杆菌 (enteropathogenic Escherichia coli)、大肠杆菌O157:′H7(E.coliO157:′H7)、土拉热弗 朗西斯菌(Francisella tularensis)、流感嗜血杆菌(Haemophilus influenzae)、幽门螺 杆菌(Helicobacter pylori)、肺炎克雷伯菌(Klebsiella pneumoniae)、嗜肺军团菌 (Legionella pneumophila)、问号钩端螺旋体(Leptospira interrogans)、单核细胞增生 性李斯特氏菌(Listeria monocytogenes)、麻风分枝杆菌(Mycobacterium leprae)、结核 分枝杆菌(Mycobacterium tuberculosis)、肺炎支原体(Mycoplasma pneumoniae)、淋病奈 瑟氏菌(Neisseria gonorrhoeae)、脑膜炎奈瑟氏菌(Neisseria meningitidis)、变形杆菌 属(Proteus)、铜绿假单胞菌(Pseudomonas aeruginosa)、立氏立克次体(Rickettsia rickettsii)、伤寒沙门氏菌(Salmonella typhi)、鼠伤寒沙门氏菌(Salmonella typhimurium)、宋内志贺氏菌(Shigella sonnei)、金黄色葡萄球菌(Staphylococcus aureus)、表皮葡萄球菌(Staphylococcus epidermis)、腐生葡萄球菌(Staphylococcus saprophytics)、耐甲氧西林金黄色葡萄球菌(methicillin-resistant Staphylococcus aureus ,MRSA)、耐万古霉素金黄色葡萄球菌(vancomycin-resistant Staphylococcus aureus ,VSA)、无乳链球菌(Streptococcus agalactiae)、肺炎链球菌(Streptococcus pneumoniae)、化脓性链球菌(Streptococcus pyogenes)、苍白密螺旋体(Treponema pallidum)、霍乱弧菌(Vibrio cholerae)和鼠疫耶尔森氏菌(Yersinia pestis)组成的组。 在另一实施方案中,细菌选自由肠球菌属(Enterococcus sp .)、埃希氏菌属 (Escherichia sp.)、葡萄球菌属(Staphylococcus sp.)、克雷伯菌属(Klebsiella sp.)、 9 CN 111601897 A 说 明 书 7/43 页 不动杆菌属(Acinetobacter sp .)、假单胞菌属(Pseudomonas sp .)和肠杆菌属 (Enterobacter sp.)组成的组。 在另一实施方案中,提供了一种用于确定表1-3任一中列出的细菌对测试抗微生 物剂的易感性的方法,该方法包括(a)在存在或不存在抗微生物剂的情况下培养表1-3任一 中列出的细菌以产生多个原代培养物;(b)在存在或不存在表1-3任一中所列的转化噬菌体 的情况下培养(a)的原代培养物,其中噬菌体对细菌是特异性的并包含用于表达异源标志 物的序列,从而产生多个次级培养物,其中第一转化次级培养物包括已经用测试抗微生物 剂处理的转化细菌和第二转化次级培养物包括未用抗微生物剂处理的转化细菌;和(c)检 测第一和第二转化次级培养物各自的标志物的水平或活性,从而确定细菌对抗微生物剂的 易感性。在该实施方案中,细菌可选自由炭疽芽孢杆菌(Bacillus anthracis)、枯草芽孢杆 菌(Bacillus subtilis)、苏云金芽孢杆菌(Bacillus thuringiensis)、大肠杆菌、德氏乳 杆菌(Lactobacillus delbrueckii)、植物乳杆菌(Lactobacillus plantarum)、乳酸乳球 菌(Lactococcus lactis)、单核细胞增生性李斯特氏菌、铜绿假单胞菌、丁香假单胞菌 (Pseudomonas syringae)、克雷伯氏菌属(Klebsiella)、沙门氏菌属(Salmonella)、志贺氏 菌属(Shigella)和金黄色葡萄球菌组成的组。 在另一实施方案中,提供了一种用于确定细菌对测试抗微生物剂的易感性的方 法。该方法包括(a)在抗微生物剂存在和/或不存在的情况下培养细菌以产生多个原代培养 物;(b)在存在或不存在转化重组或工程化噬菌体的情况下培养(a)的原代培养物,所述噬 菌体对细菌是特异性的并且包含异源标志物,从而产生多个次级培养物,其中第一次转化 次级培养物包括已经用测试抗微生物剂处理的转化细菌和第二转化次级培养物包括未用 抗微生物剂处理的转化细菌;和(c)检测第一和第二转化次级培养物各自的标志物的水平 或活性,从而确定细菌对抗微生物剂的易感性。 在一个实施方案中,重组或工程化噬菌体可选自由(a)裂解或增殖性噬菌体;(b) 温和或溶原性噬菌体和(c)丝状噬菌体组成的组。 在一个具体实施方案中,重组或工程化噬菌体是选自由T4、T7、T3和MS2组成的组 的裂解或增殖性噬菌体。在第二个具体实施方案中,重组或工程化噬菌体是温和或溶原性λ 噬菌体。在第三个具体实施方案中,重组或工程化噬菌体是选自由fl、fd和M13组成的组的 丝状噬菌体。该方法可以使用各种噬菌体的组合来实施。 在另一实施方案中,提供了一种确定细菌对测试抗微生物剂的易感性的方法。该 方法包括(a)在抗微生物剂存在和/或不存在的情况下培养细菌以产生多个原代培养物; (b)在存在或不存在对细菌特异的且包含可在细菌细胞中表达为核酸或多肽产物的标志物 的转化噬菌体的情况下培养(a)的原代培养物,从而产生多个次级培养物,其中第一转化次 级培养物包括已经用测试抗微生物剂处理的转化细菌和第二转化次级培养物包括未用抗 微生物剂处理的转化细菌;和(c)检测第一和第二转化次级培养物各自的标志物的水平或 活性,从而确定细菌对抗微生物剂的易感性。在具体实施方案中,标志物可表达为选自由抗 原、酶、抗体或其片段和适体,或其组合组成的组的多肽。 在一些实施方案中,表达的多肽标志物可包含可检测的标记。在其他实施方案中, 多肽标志物可以用选自荧光分析、化学发光分析、酶分析、凝胶电泳、免疫分析、配体结合分 析、色谱分析、光谱学或其组合的分析法进行检测。特别地,表达的多肽标志物用酶联免疫 10 CN 111601897 A 说 明 书 8/43 页 吸附分析(ELISA)或侧流免疫分析检测。在某些实施方案中,所述方法可以进一步包括通过 检测作为选自由DNA、RNA或其组合组成的组的核酸的第二标志物验证检测结果。在其中初 始检测被验证的此类实施方案中,可以用凝胶电泳、核酸扩增技术如聚合酶链反应(PCR)、 定量聚合酶链式反应(qPCR)或其组合来检测第二核酸标志物。 在另一实施方案中,提供了一种用于确定细菌对测试抗微生物剂的易感性的方 法,包括(a)在存在或不存在抗微生物剂的情况下培养细菌以产生多个原代培养物;(b)在 存在或不存在对细菌特异性且包含编码异源蛋白质的核酸的转化噬菌体的情况下培养(a) 的原代培养物,所述异源蛋白质是(1)与抗体特异性结合的抗原或(2)催化反应的酶,从而 产生多个次级培养物,其中第一转化次级培养物包括已经用测试抗微生物剂处理的转化细 菌和第二转化次级培养物包括未用抗微生物剂处理的转化细菌;和(c)检测第一和第二转 化次级培养物各自的标志物的水平或活性,从而确定细菌对抗微生物剂的易感性。在这一 实施方案下,其中异源蛋白质是(1)与抗体特异性结合的抗原,检测步骤包括用免疫分析检 测蛋白质的水平。仍然在这一实施方案下,其中异源蛋白质是(2)催化反应的酶,检测步骤 包括用酶分析检测蛋白质的活性。 在相关的实施方案中,提供了一种用于筛选测试剂对目标细菌样本的抗菌活性的 方法,包括(a)在存在或不存在测试剂的情况下培养目标细菌以产生多个原代培养物;(b) 在存在或不存在对细菌特异性且包含标志物的转化噬菌体的情况下培养(a)的原代培养 物,从而产生多个次级培养物,其中第一转化次级培养物包括已经用测试剂处理的转化细 菌和第二转化次级培养物包括未用测试剂处理的转化细菌;和(c)检测第一和第二转化次 级培养物各自的标志物的水平或活性,其中第一转化次级培养物中标志物的水平或活性与 第二转化次级培养(对照)中标志物的水平或活性相比的降低指示该测试剂具有抗菌活性。 另一实施方案是用于确定食品样品中抗生素剂的存在或不存在的方法,包括(a) 在多种细菌培养物中培养食品样品,其中第一培养物包含对抗生素敏感的细菌和第二培养 物包含对抗生素抗性的细菌,从而产生多个原代培养物;(b)在存在或不存在对细菌特异性 且包含标志物的转化噬菌体的情况下培养(a)的原代培养物,从而产生多个次级培养物,其 中第一转化次级培养物包括对抗生素剂敏感的转化细菌和第二转化次级培养物包括对抗 生素剂抗性的转化细菌;和(c)检测第一和第二转化次级培养物各自的标志物的水平或活 性,其中第一转化次级培养物中标志物的水平或活性与第二转化次级培养物(对照)中标志 物的水平或活性相比的降低指示食物样品包含抗生素剂。在该实施方案中,易感细菌和抗 性细菌属于相同菌株。此外,抗性细菌可以是易感细菌的突变变体,其包含赋予对抗细菌剂 的抗性的重组基因。 另一实施方案涉及用于测定抗细菌剂对目标细菌样品的最小抑制浓度(MIC)的方 法,包括(a)在存在或不存在不同浓度的抗细菌剂的情况下培养目标细菌以产生多个原代 培养物;(b)在存在或不存在对细菌特异性且包含标志物的转化噬菌体的情况下培养(a)的 原代培养物,从而产生多个次级培养物,其中实验组包括已经用各种浓度的抗细菌剂处理 的转化细菌和对照组包括未用测试剂处理的转化细菌;和(c)检测实验组和对照组各自的 标志物的水平或活性,其中抗细菌剂能够与对照组中的阈值水平或活性相比降低标志物的 水平或活性的最小浓度是MIC的表示。在相关的实施方案中,提供了用于确定两种或更多种 抗细菌剂的加性、超加性、协同或拮抗活性的方法,该方法包括根据前述确定每种抗细菌剂 11 CN 111601897 A 说 明 书 9/43 页 的最小抑制浓度(MIC)和根据前述确定包含最小抑制浓度的每种药剂的组合的抑制作用; 比较该组合的抑制作用与单一药剂的抑制作用,从而确定两种或多种抗细菌剂的加性、超 加性、协同或拮抗活性。 又一个实施方案涉及在需要的受试者中诊断和治疗细菌性疾病的方法,包括(a) 培养包含细菌的多个受试者样品以产生多个原代细菌培养物;(b)在多种转化噬菌体的存 在下培养(a)的原代培养物,其中每种转化噬菌体对细菌是特异性的并且包含编码独特多 肽标志物的核酸,从而产生多个次级培养物;(c)通过免疫检测来检测次级培养物中的独特 多肽标志物;(d)将标志物的检测与细菌相关联;(e)将细菌的存在与细菌疾病相关联;和 (f)任选地向受试者施用对检测的细菌特异性的抗生素剂,从而治疗细菌性疾病。 附图说明 在附图/表格和以下描述中阐述了本发明的一个或多个实施方案的细节。根据附 图/表格和详细说明以及从权利要求,本发明的其他特征、目的和优点将是清楚的。 图1示出了根据本文描述的方法的一个实施方案的示例性工作流程;和 图2示出了根据本文描述的方法的另一实施方案的示例性工作流程。 详细说明 本文描述的实施方案提供了使用重组噬菌体诊断或检测细菌感染因子和疾病的 方法和分析。该方法适用于检测细菌感染因子以及适用于确定这些感染因子的药物抗性。 此外,该方法用于提供关于感染因子对抗微生物剂的易感性的信息。 A.感染菌 基本上任何细菌可以被检测,并且该方法和组合物可用于确定细菌的抗生素敏感 性或用于筛选对目标细菌施加所需(例如抗微生物或细胞毒性)作用的候选抗生素剂。 在一个实施方案中,细菌是革兰氏阴性细菌。典型的革兰氏阴性细菌包括变形菌 如大肠杆菌、沙门氏菌、假单胞菌和螺杆菌,以及蓝细菌。当与药物结合分类时,它们包括引 起呼吸系统紊乱的铜绿假单胞菌和流感嗜血杆菌,引起泌尿系统紊乱的大肠杆菌和奇异变 形杆菌(Proteus mirabilis),以及引起消化系统紊乱的幽门螺杆菌和格特纳杆菌 (Bacillus Gaertner) ,及微球菌如脑膜炎奈瑟氏菌、卡他莫拉菌(Moraxella catarrhalis)和淋病奈瑟氏菌。 在另一实施方案中,细菌是革兰氏阳性细菌。“革兰氏阳性细菌”是指在其细胞壁 中含有磷壁酸(例如,脂磷壁酸和/或壁磷壁酸)或功能等同的含糖聚合物(例如,鼠李多糖、 糖醛酸磷壁酸、阿拉伯半乳聚糖、脂甘露聚糖和脂阿拉伯甘露聚糖)的一种或多种细菌。功 能等同的含糖聚合物的非限制性实例描述于Weidenmaier等,Nature,6:276-287,2008中。 功能等同的含糖聚合物的其他实例是本领域已知的。在一些实施方案中,使用革兰氏染色 法鉴定革兰氏阳性细菌(例如,通常包括用结晶紫染色,用碘溶液处理,用酒精脱色和用番 红复染的步骤,其中革兰氏阳性细菌保留紫色染色)。本文描述了革兰氏阳性细菌的非限制 性实例。革兰氏阳性细菌的其他实例是本领域已知的。本文描述了用于检测或鉴定革兰氏 阳性细菌的示例性方法。用于检测或鉴定革兰氏阳性细菌的其他方法是本领域已知的。 目标细菌包括感染哺乳动物宿主(例如牛科动物、鼠科动物、马科动物、灵长类动 物、猫科动物、犬科动物和人宿主)的病原菌。在一个实施方案中,细菌感染人类宿主和/或 12 CN 111601897 A 说 明 书 10/43 页 引起人类宿主的疾病。这种病原细菌的实例包括例如拟杆菌属(Bacteroides)、梭菌属 (Clostridium)、链球菌属(Streptococcus)、葡萄球菌属(Staphylococcus)、假单胞菌属 (Pseudomonas)、嗜血杆菌属(Haemophilus)、军团菌属(Legionella)、分枝杆菌属 (Mycobacterium)、埃希氏菌属(Escherichia)、沙门氏菌属(Salmonella)、志贺氏菌属 (Shigella)、弧菌属(Vibrio)或李斯特菌属(Listeria)的细菌物种的成员。在人类宿主中 引起疾病的病原细菌的一些临床相关实例包括但不限于炭疽芽孢杆菌、蜡状芽孢杆菌、百 日咳博德特氏菌、伯氏疏螺旋体、流产布鲁氏菌、犬布鲁氏菌、羊布鲁氏菌、猪布鲁氏菌、空 肠弯曲杆菌、肺炎衣原体、鹦鹉热衣原体、沙眼衣原体、肉毒梭菌、艰难梭菌、产气荚膜梭菌、 破伤风梭菌、白喉棒状杆菌、粪肠球菌、耐万古霉素粪肠球菌、屎肠球菌、大肠埃希菌、产肠 毒素大肠杆菌(ETEC)、肠致病性大肠杆菌、大肠杆菌0157:H7、土拉热弗朗西斯菌、流感嗜血 杆菌、幽门螺杆菌、嗜肺军团菌、问号钩端螺旋体、单核细胞增生性李斯特氏菌、麻风分枝杆 菌、结核分枝杆菌、肺炎支原体、淋病奈瑟氏菌、脑膜炎奈瑟氏菌、变形杆菌属、铜绿假单胞 菌、立氏立克次体、伤寒沙门氏菌、鼠伤寒沙门氏菌、宋内志贺菌、金黄色葡萄球菌、表皮葡 萄球菌、腐生葡萄球菌、耐甲氧西林金黄色葡萄球菌(MRSA)、耐万古霉素金黄色葡萄球菌 (VSA)、无乳链球菌、肺炎链球菌、化脓性链球菌、苍白密螺旋体、霍乱弧菌和鼠疫耶尔森氏 菌。 在另一实施方案中,感染细菌选自由艰难梭菌、耐碳青霉烯肠杆菌科(CR- Klebsiella spp;CR-E.coli)和淋病奈瑟氏菌组成的组。在另一实施方案中,感染细菌选自 由耐多药不动杆菌、耐药性弯曲杆菌、产超广谱β-内酰胺酶(ESBL)肠杆菌科、耐万古霉素肠 球菌、耐多药铜绿假单胞菌、耐药性非伤寒沙门氏菌、耐药性肠道沙门氏菌伤寒血清型、耐 药性志贺氏菌、耐甲氧西林金黄色葡萄球菌(MRSA)、耐药性肺炎链球菌和耐药性结核菌组 成的组。在另一实施方案中,感染细菌选自由耐万古霉素金黄色葡萄球菌、耐红霉素A群链 球菌、耐克林霉素B群链球菌组成的组。 在某些实施方案中,感染因子天然存在于宿主受试者中。在另一实施方案中,感染 因子是宿主受试者外源的侵入性物种。优选地,宿主是哺乳动物,例如,啮齿动物、人类、牲 畜、伴侣动物或者非驯养或野生动物。在一个实施方案中,受试者可以是啮齿动物,例如,小 鼠、大鼠、豚鼠等。在另一实施方案中,受试者可以是牲畜。合适的牲畜的非限制性实例可包 括猪、牛、马、山羊、绵羊、美洲驼和羊驼。在又一个实施方案中,受试者可以是伴侣动物。伴 侣动物的非限制性实例可包括宠物,例如狗、猫、兔和鸟。在又一个实施方案中,受试者可以 是动物园动物。如本文所用,“动物园动物”是指可以在动物园中发现的动物。这些动物可包 括非人灵长类动物、大型猫、狼和熊。在示例性实施方案中,受试者是人。 该方法可用于分析各种样品中包含的感染因子,所述样品包括例如生物样品、研 究测试样品、环境样品(例如水样,包括选自天然水体、池塘、社区水库、休闲水域、游泳池、 漩涡浴池、热水浴池、水疗中心、水上公园、天然淡水和海洋表面水域的水样)和工业样品 (如发酵接种物(如乳酸菌)、化学试剂、培养基、清洁液)。 优选地,样品是包含体液的生物样品,例如痰、泪液、唾液、汗液、粘液、血清、精液、 尿液、粪便、呕吐物和血液。样品可以包括例如脑脊髓液(CSF)、血浆、血清、淋巴液、肺灌洗 液、胸膜液等。在一些实施方案中,样品可以使用任何已知的装置或方法从受试者获得,例 如,拭子、尿道导管、抽吸器、皮下注射针、细针活组织检查、空心针活组织检查、钻孔活组织 13 CN 111601897 A 说 明 书 11/43 页 检查、代谢笼和注射器。 在一些实施方案中,生物样品进行处理以用于本文所述的方法。作为非限制性实 例,将痰液或气道表面液(airway surface fluid)(ASF)收集在合适的容器中,例如无菌样 品瓶。样品例如使用乙腈溶解至约60%的终浓度,使用三氟乙酸溶解至约0.1%的终浓度, 或使用N-乙酰半胱氨酸。 在某些实施方案中,可以操纵生物样品以培养其中包含的细菌。术语“培养物”是 指培养的细胞、培养物上清液、其混合物或培养滤液(如果使用液体培养基);如果使用固体 培养基,术语“培养物”是指细胞和它们在其上生长的培养基的混合物。例如,如果使用液体 培养基,可以通过以下过程从培养混合物中回收标志物。当达到细菌的完全生长时,培养混 合物进行抗生素和/或噬菌体的处理。这种下游处理可以插入一个或多个包括离心或过滤 的洗涤和/或分离步骤,以获得不含污染物的粗制细菌制剂。标志物可以在细胞水平(例如, 原位)或在使培养物经受进一步处理之后检测或分析。例如,其中标志物是细胞溶质中的蛋 白质或DNA,它们可以通过使用合适的方法例如研磨或超声处理破坏细胞来提取。细胞可以 在培养基中直接进行超声处理以破坏细胞,并且粗酶溶液可以通过从处理的溶液除去任何 不溶物质来获得。 如果在固体培养基上进行培养,标志物可以通过首先使用以下过程操作培养物来 分析:将水加入到含有培养细胞的固体培养基中,且任何不溶物质立即或在通过合适的方 法如超声处理破坏细胞后从混合物中除去。粗标志物制剂可以通过常规纯化技术从粗裂解 物分离,例如有机溶剂分馏、硫酸铵分级分离、透析、等电沉淀和柱色谱,它们可以单独或组 合使用。标志物的水平或活性可以使用常规方法测定,例如抗原蛋白质标志物的免疫分析、 抗体样标志物的配体结合、酶样标志物的酶分析、核酸杂交和/或核酸扩增等。 根据目的,可以使用常规技术分析细胞培养物。例如,可以将细菌培养至对数期 (MSSA USA300和MRSA USA300),并且可以使用常规技术例如分光光度法检测对数期峰值的 噬菌体(peak logarithmic phage)。使用对数期细菌可能是优选的,因为它们由于粘附素 的较高表达而更可能是粘附的,并且与稳定期细胞相比,它们的肽聚糖层可能较少交联和 是厚的和细胞具有更高代谢活性,从而更快地对损害作出响应。然而,最佳条件可能因菌株 而异。由于在临床环境中经常遇到不同的菌株,因此该信息对于评估诊断方法的效用是重 要的。尽管本文设想存在菌株变异性,但预期细菌的表现足够相似以允许使用单一方案来 测试所有菌株。这种预期是基于以下事实:细菌家族(例如,葡萄球菌)在遗传上彼此非常相 似,且因此具有相似的细胞结构,这将是它们对特定噬菌体的响应性的主要成分。 在一些实施方案中,该方法和组合物可用于测定微生物例如细菌的易感性。如本 文所用,术语“易感性”是指细菌细胞受抗生素影响的程度。也就是说,细胞可能根本不受影 响,它可能会使其生长和增殖减慢或停止而不被杀死,或者可能被杀死。易感性还指细菌物 种或菌株的群体受抗生素影响的程度。在这种情况下,群体的某些高度易感的细胞可能非 常敏感,并且可能被非常低浓度的抗生素杀死,其他不太敏感的细胞可能使其生长和增殖 减慢,而其他可能根本不受影响。 在相关的实施方案中,所述方法和组合物可用于鉴定微生物例如细菌对抗微生物 剂或抗生素的抗性。本文中的术语“对抗生素的抗性”是指特定的细菌菌株(通常是突变菌 株)不被杀死,或与衍生该菌株的相应野生型菌株相比,显著更慢地被杀死。抗性还可以通 14 CN 111601897 A 说 明 书 12/43 页 过突变和野生型菌株的改变的生长特性来反映。例如,培养基中低浓度的抗生素阻止或显 著降低野生型菌株的生长,同时突变菌株的生长不受影响。抗性菌株的表型,例如,改变的 生长、细胞分裂、代谢、生物膜产生、毒力等,可以使用常规技术来确定,例如,在相同条件下 生长野生型和突变菌株以评估测量的参数中的变化。敏感菌株可用作抗性评估的参考标准 (阳性对照)。 在一个实施方案中,该方法通过在存在和不存在抗生素的情况下培养细菌样品来 进行。可以改变或调整培养基或发酵培养基以满足相应菌株的需要。用于各种微生物的培 养基的描述存在于美国细菌学会的“Manual of Methods for General Bacteriology” (Washington D.C.,USA,1981)中。术语培养基和发酵培养基或培养基是可互换的。 从其最简单的意义上,培养基含有至少一种碳源(例如葡萄糖)和至少一种氮源 (例如硝酸盐),任选地还含有磷源,例如磷酸、磷酸钾或其他磷酸盐。优选地,将培养基缓冲 用于细菌生长。培养基可另外包含盐,例如金属(例如钠、钾、镁、钙和铁)的氯化物或硫酸 盐,例如硫酸镁或硫酸铁,其促进生长和/或代谢活性。最后,根据需要,可以向培养基中加 入必需的生长因子,如氨基酸,例如高丝氨酸,和维生素,例如硫胺素、生物素或泛酸。参见 美国专利No.9,074,229。 将含有细菌的起始样品以单批的形式加入培养物中或在培养期间以合适的方式 加料,例如每2-4小时或每1-3小时,或者每1、2、3或4小时。 培养物的pH可以通过以合适的方式使用碱性化合物如氢氧化钠、氢氧化钾、氨或 氨水或者酸性化合物如磷酸或硫酸来控制。通常将pH调节至6.0至8.5,优选6.5至8。为了控 制发泡,可以使用消泡剂,例如聚乙二醇脂肪酸酯。为了保持细菌的稳定性,可能向培养基 中加入合适的选择性物质,例如诱导剂如IPTG。发酵优选在有氧条件下进行。为了维持这些 条件,将氧或含氧气体混合物(例如空气)引入培养物中。在分批或流加方法中,优选持续培 养直至达到微生物的所需的密度的量。检测通过分光光度法(吸收、荧光)方式进行。这个目 标通常在2小时到160小时内完成。在连续方法中,较长的培养时间是可能的。微生物的活性 导致在发酵培养基中和/或微生物细胞中各种标志物的浓缩(累积)。 合适的发酵培养基的实例可以特别地在美国专利No.5,770 ,409、5,275 ,940、5, 827,698、5,756,345;及WO 2007/012078和WO 2009/043803。 B.抗生素 上述培养基可以补充或不补充抗生素。如本文所用,术语“抗生素”或“抗微生物 剂”是指抑制微生物生长或破坏微生物的物质。优选地,抗生素可用于控制感染因子的毒力 和/或治疗传染病。抗生素还指半合成物质,其中由微生物例如酵母或真菌产生的天然形式 随后在结构上被修饰。 在另一实施方案中,培养基可以补充或不补充益菌物质。如本文所用,术语“益菌” 是指促进微生物生长或代谢活性的物质,例如,微量营养素、生长诱导物质或去除毒素的物 质。 优选地,抗生素选自由β-内酰胺类(包括β-内酰胺酶抑制剂和头孢菌素类)、氟喹 诺酮类、氨基糖苷类、四环素类和/或甘氨酰环素类和/或多粘菌素类组成的组。还可以测试 抗微生物剂的任何组合,例如,至少一种β-内酰胺和至少一种氟喹诺酮;至少一种氨基糖苷 类和一种头孢菌素;至少一种β-内酰胺和一种β-内酰胺酶抑制剂,任选地与一种氨基糖苷 15 CN 111601897 A 说 明 书 13/43 页 类一起,等。 如本文所用,术语“β-内酰胺”是指在其分子结构中含有β-内酰胺环的任何抗生素 剂。代表性实例包括天然和半合成青霉素类和青霉素衍生物、克拉维酸、碳青霉烯类、头孢 菌素类、头霉素类和单内酰环类。这些药物被广泛称为“β-内酰胺酶”的酶代谢。β-内酰胺酶 分为四个分子类别(A、B、C和D)。A类酶优先水解青霉素类;B类酶包括具有比其他酶更宽的 底物谱的金属酶;C类酶负责革兰氏阴性菌对多种抗生素的抗性;和D类酶是丝氨酸水解酶, 其表现出独特的底物谱。 通常,β-内酰胺根据其核心环结构分类和分组,其中每个组可以分成不同的类别。 术语“培南(penam)”用于描述青霉素抗生素的成员的核心骨架,例如含有噻唑烷环的β-内 酰胺类。青霉素类可包括窄谱青霉素类,例如苄星青霉素、苄青霉素(青霉素G)、苯氧甲基青 霉素(青霉素V)、普鲁卡因青霉素和苯唑西林。窄谱耐青霉素酶抗性青霉素类,如甲氧西林、 双氯西林和氟氯西林。窄谱耐β-内酰胺酶青霉素可包括替莫西林。中谱青霉素类包括例如 阿莫西林和氨苄青霉素。广谱青霉素类包括阿莫克拉(阿莫西林 克拉维酸)。最后,青霉素 组还包括超广谱青霉素类,例如,阿洛西林、羧苄青霉素、替卡西林、美洛西林和哌拉西林。 合成青霉素衍生物包括例如法罗培南。 含有吡咯烷环的β-内酰胺被称为碳青霉烷类(carbapenams)。碳青霉烯组包括:比 阿培南、多利培南、厄他培南、亚胺培南、美罗培南、帕尼培南和PZ-601。 头孢菌素类和头霉素类包括头孢氨苄、头孢噻吩、头孢唑啉、头孢克洛、头孢呋辛、 头孢孟多、头孢替坦、头孢西丁、头孢噻肟和头孢泊肟。对革兰氏阳性菌有活性的第四代头 孢菌素包括头孢吡肟和头孢匹罗。头孢菌素类可进一步包括:头孢羟氨苄、头孢克肟、头孢 丙烯、头孢氨苄、头孢噻吩、头孢呋辛、头孢孟多、头孢吡肟和头孢匹罗。头霉素类包括例如 头孢西丁,头孢替坦,头孢美唑和氟氧头孢。 碳头孢烯类的一个例子是氯碳头孢。对革兰氏阴性细菌活性的单内酰环类包括, 例如,替吉莫南、诺卡霉素A和野火菌毒素。合成的头孢烯类包括,例如,克拉维酸和氧头孢 烯类如拉氧头孢和氟氧头孢。 氟喹诺酮类通过抑制对于细菌DNA复制必需的酶起作用。其代表性实例包括环丙 沙星、加雷沙星、加替沙星、吉米沙星、左氧氟沙星和莫西沙星。 氨基糖苷类对大多数革兰氏阴性需氧和兼性厌氧芽孢杆菌具有杀菌活性。代表性 实例包括,例如,卡那霉素、阿米卡星、妥布霉素、地贝卡星、庆大霉素、西索米星、奈替米星、 新霉素B、新霉素C、新霉素E(巴龙霉素)和链霉素,包括合成衍生物克拉霉素和阿奇霉素。 四环素类是具有八氢并四苯-2-甲酰胺骨架的聚酮化合物的亚类。它们可以是天 然存在的(例如,四环素、金霉素、土霉素、地美环素)或半合成的(例如,赖甲环素、甲氯环 素、美他环素、米诺环素、罗利环素)。甘氨酰环素类(例如米诺环素/替加环素)衍生自四环 素类。 多粘菌素类是对革兰氏阴性细菌如大肠杆菌和铜绿假单胞菌具有活性的多肽抗 生素。临床上仅使用多粘菌素B和多粘菌素E(粘菌素)。 在实施该方法时,培养基可以补充上述抗生素中的一种或多种。抗生素的浓度可 以根据抗生素和测试的菌株类型而变化。优选地,抗生素的剂量等于或大于特定抗生素对 特定菌株的最小抑制浓度(MIC)。用于确定MIC的方法是本领域已知的(参见,Andrews等, 16 CN 111601897 A 说 明 书 14/43 页 JAntimicrob Chemother.,48 Suppl 1:5-16,2001)。40种左右的抗微生物剂针对四种细菌 菌株(大肠杆菌、金黄色葡萄球菌、铜绿假单胞菌和粪肠球菌)的MIC的代表性图表显示在欧 盟药敏试验标准委员会(European Committee for Antimicrobial Susceptibility Testing,EUCAST)发表的题为"Determination of minimum inhibitory concentrations (MICs)of antibacterial agents by broth dilution"的报告(European Society of Clinical Microbiology and Infectious Diseases CMI,9,1-7,2003)的表3中。 通常,可以提高抗生素的浓度以鉴定或检测抗性菌株,例如,提高到基线MIC的至 少2倍、至少5倍、至少10倍、至少20倍、至少50倍、至少100倍、至少300倍或甚至1000倍。这在 目标细菌和抗生素对细菌的MIC已知的情况中特别有效。例如,对于大肠杆菌,大多数抗生 素的MIC范围可以为约0.01mg/L至约10mg/L;然而,抗性菌株可能直到浓度提高到例如基线 水平上10倍(即,1log倍)-1000倍(即,3-log倍)才是易感的。在这一方面,可以相应地调整 最终的抗生素浓度。 纯粹为了说明的目的,可以使用以下剂量-用于测试细菌对β-内酰胺类如阿莫西 林的抗性,浓度范围可以为约2mg/L-约40mg/L,特别是约5mg/L-约20mg/L。参见美国专利 No.9,347,888。另一方面,为了测试细菌对氯唑西林的抗性,浓度范围可以在约25mg/L和约 300mg/L之间。对于碳青霉烯,浓度范围可以在0.05和32mg/L之间。这包括对于法罗培南的 约2mg/L至约32mg/L和对于多利培南的约0.05mg/L至约2mg/L的范围(参见Woodman等,J Med Microbiol .,19(1):15-23 ,1983)。对于头孢菌素类,浓度范围可以为约1mg/L至约 20mg/L,优选约4mg/L至约16mg/L(参见Waterworth,J Clin Pathol,35:1177-1180,1982)。 更特别地,抗生素可以以0.1mg/mL、0.5mg/L、1mg/mL、2mg/mL、3mg/mL、4mg/mL、 5mg/mL、6mg/mL、7mg/mL、8mg/mL、9mg/mL、10mg/mL、11mg/mL、12mg/mL、13mg/mL、14mg/mL、 15mg/mL、16mg/mL、17mg/mL、18mg/mL、19mg/mL、20mg/mL、21mg/mL、22mg/mL、23mg/mL、24mg/ mL、25mg/mL、26mg/mL、27mg/mL、28mg/mL、29mg/mL、30mg/mL、31mg/mL、32mg/mL、33mg/mL、 34mg/mL、35mg/mL、36mg/mL、37mg/mL、38mg/mL、39mg/mL、40mg/mL、41mg/mL、42mg/mL、43mg/ mL 44mg/mL、45mg/mL、50mg/mL、60mg/mL、70mg/mL、80mg/m、90mg/mL、100mg/mL、150mg/mL、 200mg/mL、250mg/mL、300mg/mL、400mg/mL、500mg/mL或更高中任一的浓度使用。例如,亚胺 培南和厄他培南可以以50、30、20、15、10、5和1mg/mL的浓度使用。剂量对于抗生素的组合可 以类似地调整,例如,通过首先对于野生型菌株确定MIC(复合药剂)和逐步提高剂量以鉴定 抗性菌株。 细菌在存在或不存在抗生素的情况下培养特定的时间段,例如2小时至160小时, 特别是8小时至24小时,特别是10小时至16小时。在与噬菌体接触之前,细菌可处于其生长 期或静止期。生长期是以细胞倍增为特征的时期,其中培养物中细胞的数量呈指数增长。静 止期是由新细菌的生长和衰老细胞的死亡两者引起的,这通常是由于生长限制因素如必需 营养物的耗尽或废物的积累。优选地,在接种噬菌体之前,细菌处于生长期。用于确定细菌 生长期的方法是本领域已知的。参见Hall等,Mol Biol Evol,31(l):232-8,2014。 在一个实施方案中,在接种噬菌体之前,细菌用抗生素处理。原代培养物可以在接 种前任选地洗涤,例如,用洗涤缓冲液。取决于存活培养物的密度,原代培养物或其洗涤沉 淀(在原代培养物离心后获得)可以在已经接种噬菌体的新鲜原始培养基(或含抗生素的培 养基)中再生长。 17 CN 111601897 A 说 明 书 15/43 页 在另一实施方案中,细菌在用抗生素剂处理的同时用噬菌体接种。该实施方案可 能特别适合于非裂解噬菌体。 C.噬菌体 本方法的实施方案利用宿主特异性的噬菌体。如本文所用,术语“噬菌体”具有本 领域所理解的常规含义,例如选择性感染一种或多种细菌的病毒。许多噬菌体对于特定细 菌属或种或菌株是特异性的。术语“噬菌体(bacteriophage)”与术语“噬菌体(phage)”同 义。噬菌体可包括但不限于属于以下病毒科任一种的噬菌体 :覆盖噬菌体科 (Corticoviridae)、囊状噬菌体科(Cystoviridae)、丝状噬菌体科(Inoviridae)、光滑噬菌 体科(Leviviridae)、微小噬菌体科(Microviridae)、肌尾噬菌体科(Myoviridae)、短尾噬 菌体科(Podoviridae)、长尾噬菌体科(Siphoviridae)或复层噬菌体科(Tectiviridae)。噬 菌体可以是裂解性噬菌体或溶原性噬菌体或丝状噬菌体。裂解性噬菌体是遵循通过完成裂 解周期的裂解途径而不是进入溶原途径的噬菌体。裂解性噬菌体经历病毒复制,其导致细 胞膜裂解、细胞破坏以及能够感染其他细胞的后代噬菌体颗粒的释放。溶原性噬菌体是能 够进入溶原途径的噬菌体,其中噬菌体在其裂解周期完成之前成为细胞基因组的休眠、被 动部分。丝状噬菌体包含包装成长丝的环状单链脱氧核糖核酸(ssDNA)基因组。这些噬菌体 不通过裂解细菌繁殖;相反,它们被分泌到环境中而不杀死宿主。 在一个实施方案中,噬菌体是裂解或增殖性噬菌体(例如,T4、T7、T3和MS2)。在另 一实施方案中,噬菌体是温和或溶原性噬菌体(例如,λ噬菌体)。在又一实施方案中,噬菌体 是丝状噬菌体(例如,fl、fd和M13)。也可以使用各种噬菌体的组合。噬菌体展示技术是本领 域已知的,例如美国专利号8,685 ,893;美国专利号7,811 ,973;和美国专利公开号2002- 0044922。优选地,噬菌体能够转化宿主细菌。如本文所用,术语“转化”是指将DNA引入宿主 细胞中,使得DNA可以作为染色体外元件或通过染色体整合复制。也就是说,转化是指通过 将外源DNA引入细胞中的基因合成改变。如本领域中所公认的,大多数细菌的DNA包含在称 为细菌染色体的单一环状分子和一种或多种质粒中。 噬菌体是工程化或重组噬菌体,其用作对于天然噬菌体外源的基因的载体。如本 文所用,术语“重组噬菌体”或“工程化噬菌体”是含有非天然存在的核酸序列或具有通过人 工组合两个另外分开的序列区段形成的序列的噬菌体。这种人工组合可以通过化学合成或 分离的核酸片段的人工操作来完成,例如,通过基因工程技术或使用DNA转座。类似地,重组 蛋白质是由重组核酸分子编码的蛋白质。术语重组噬菌体包括仅通过插入核酸而改变的噬 菌体,例如通过插入用于编码用作报告分子或指示分子的异源蛋白质的核酸。 在某些实施方案中,噬菌体是纯化的噬菌体。术语纯化的不需要绝对纯度;相反, 它意图作为相对术语。纯化的分子是其中分子比其在天然环境中更富集的分子,例如其中 分子占到样品中类似分子的总含量的至少50%、至少60%、至少80%、至少90%、至少99% 或更高的含量。例如,重组噬菌体的纯化样品是其中重组噬菌体占样品中所有噬菌体的至 少50%的样品。 致病细菌属及其已知的宿主特异性噬菌体的列表在以下段落中给出,并且表1-3 中提供了优选类型的细菌-噬菌体对。括号中标明了同义词和拼写变体。同名词每当它们出 现时被重复(例如,D,D,d)。未命名的噬菌体在其属旁边用“NN”表示。 放线菌属的细菌被以下噬菌体感染:Av-1、Av-2、Av-3、BF307、CT1、CT2、CT3、CT4、 18 CN 111601897 A 说 明 书 16/43 页 CT6、CT7、CT8和1281。 气单胞菌属的细菌被以下噬菌体感染:AA-1、Aeh2、N、PM1、TP446、3、4、11、13、29、 31、32、37、43、43-10T、51、54、55R 1、56、56RR2、57、58、59.1、60、63、Aeh1、F、PM2、1、25、31、 40RR2.8t、(syn=44R)、(syn=44RR2.8t)、65、PM3、PM4、PM5和PM6。 芽孢杆菌属的细菌被以下噬菌体感染:A、aiz1、A1-K-I、B、BCJA1、BC1、BC2、BLL1、 BL1、BP142、BSL1、BSL2、BS1、BS3、BS8、BS15、BS18、BS22、BS26、BS28、BS31、BS104、BS105、 BS106、BTB、B1715V1、C、CK-1、Col1、Cor1、CP-53、CS-1、CS1、D、D、D、D5、ent1、FP8、FP9、PS1、 FS2、FS3、FS5、FS8、FS9、G、GH8、GT8、GV-1、GV-2、GT-4、g3、g12、g13、g14、g16、g17、g21、g23、 g24、g29、H2、ken1、KK-88、Kum1、Kyu1、J7W-1、LP52、(syn=LP-52)、L7、Mex1、MJ-1、mor2、MP- 7、MP10、MP12、MP14、MP15、Neo1、No 2、N5、N6P、PBC1、PBLA、PBP1、P2、S-a、SF2、SF6、Sha1、 Sil1、SPO2、(syn=ΦSPP1)、SΡβ、STI、ST1、SU-11、t、Tb1、Tb2、Tb5、Tb10、Tb26、Tb51、Tb53、 Tb55、Tb77、Tb97、Tb99、Tb560、Tb595、Td8、Td6、Td15、Tg1、Tg4、Tg6、Tg7、Tg9、Tg10、Tg11、 Tg13、Tg15、Tg21、Tin1、Tin7、Tin8、Tin13、Tm3、Toc1、Tog1、tol1、TP-1、TP-10vir、TP-15c、 TP-16c、TP-17c、TP-19、TP35、TP51、TP-84、Tt4、Tt6、A型、B型、C型、D型、E型、 VA-9、W、 wx23、wx26、Yun1、α、γ、p11、 1A、1B、1-97A、 1-97B、2、2、3、3、3、5、12、14、20、30、35、36、37、38、41C、51、63、64、138D、I、II、IV、NN-芽孢杆 菌(13)、ale1、AR1、AR2、AR3、AR7、AR9、Bace-11、(syn=11)、Bastille、BL1、BL2、BL3、BL4、 BL5、BL6、BL8、BL9、BP124、BS28、BS80、Ch、CP-51、CP-54、D-5、dar1、den1、DP-7、ent2、FoS1、 FoS2、FS4、FS6、FS7、G、gal1、gamma、GE1、GF-2、GS1、GT-1、GT-2、GT-3、GT-4、GT-5、GT-6、GT- 7、GV-6、g15、I9、I10、IS1、K、MP9、MP13、MP21、MP23、MP24、MP28、MP29、MP30、MP32、MP34、 MP36、MP37、MP39、MP40、MP41、MP43、MP44、MP45、MP47、MP50、NLP-1、No.1、N17、N19、PBS1、 PK1、PMB1、PMB12、PMJ1、S、SPO1、SP3、SP5、SP6、SP7、SP8、SP9、SP10、SP-15、SP50、(syn=SP- 50)、SP82、SST、sub1、SW、Tg8、Tg12、Tg13、Tg14、thu1、thu4、thu5、Tin4、Tin23、TP-13、TP33、 TP50、TSP-1、V型、VI型、V、Vx、β22、 1、1、2、2C、3NT、4、5、6、7、8、9、10、 12、12、17、18、19、21、138、III、4(巨大芽孢杆菌)、4(球形芽孢杆菌)、AR13、BPP-10、BS32、 BS107、B1、B2、GA-I、GP-10、GV-3、GV-5、g8、MP20、MP27、MP49、Nf、PP5、PP6、SF5、Tg18、TP-1、 Versailles、 1-97、837/IV、NN-芽孢杆菌(1)、Bat10、BSL10、BSL11、BS6、BS11、 BS16、BS23、BS101、BS102、g18、mor1、PBL1、SN45、thu2、thu3、Tm1、Tm2、TP-20、TP21、TP52、F 型、G型、IV型、NN-芽孢杆菌(3)、BLE、(syn=θc)、BS2、BS4、BS5、BS7、B10、B12、BS20、BS21、F、 MJ-4、PBA12、AP50、AP50-04、AP50-11、AP50-23、AP50-26、AP50-27和Bam35。以下芽孢杆菌特 异性噬菌体是缺陷型的:DLP10716、DLP-11946、DPB5、DPB12、DPB21、DPB22、DPB23、GA-2、M、 No.1M、PBLB、PBSH、PBSV、PBSW、PBSX、PBSY、PBSZ、phi、SPα、1型和μ。 拟杆菌属的细菌被以下噬菌体感染:ad12、Baf-44、Baf-48B、Baf-64、Bf-1、Bf-52、 B40-8、F1、β1、 11、67.1、67.3、68.1、NN-拟杆菌(3)、Bf42、Bf71和BF-41。 博德特氏菌属的细菌被以下噬菌体感染:134和NN-博德特氏菌(3)。 疏螺旋体属的细菌被以下噬菌体感染:NN-疏螺旋体(1)和NN-疏螺旋体。 布鲁氏菌属的细菌被以下噬菌体感染:A422、Bk、(syn=Berkeley)、BM29、FO1、 (syn=FO1)、(syn=FQ1)、D、FP2、(syn=FP2)、(syn=FD2)、Fz、(syn=Fz75/13)、(syn= Firenze 75/13)、(syn=Fi)、F1、(syn=F1)、F1m、(syn=F1m)、(syn=Fim)、F1U、(syn= 19 CN 111601897 A 说 明 书 17/43 页 F1U)、(syn=FiU)、F2、(syn=F2)、F3、(syn=F3)、F4、(syn=F4)、F5、(syn=F5)、F6、F7、 (syn=F7)、F25、(syn=F25)、(syn=f25)、F25U、(syn=F25u)、(syn=F25U)、(syn=F25V)、 F44、(syn=F44)、F45、(syn=F45)、F48、(syn=F48)、I、Im、M、MC/75、M51、(syn=M85)、P、 (syn=D)、S708、R、Tb、(syn=TB)、(syn=Tbilisi)、W、(syn=Wb)、(syn=Weybridge)、X、3、 6、7、10/1、(syn=10)、(syn=F8)、(syn=F8)、12m、24/II、(syn=24)、(syn=F9)、(syn= F9)、45/III、(syn=45)、75、84、212/XV、(syn=212)、(syn=F10)、(syn=F10)、371/XXIX、 (syn=371)、(syn=F11)、(syn=P11)和513。 伯克霍尔德氏菌属的细菌被以下噬菌体感染:CP75。 弯曲杆菌属的细菌被以下噬菌体感染:C型、NTCC12669、NTCC12670、NTCC12671、 NTCC12672、NTCC12673、NTCC12674、NTCC12675、NTCC12676、NTCC12677、NTCC12678、 NTCC12679、NTCC12680、NTCC12681、NTCC12682、NTCC12683、NTCC12684、32f、111c、191、Vfi- 6、(syn=V19)、VfV-3、V2、V3、V8、V16、(syn=Vfi-1)、V19、V20(V45)、V45、(syn=V-45)和 NN-弯曲杆菌(1)。 衣原体属的细菌被以下噬菌体感染:Chp1。 梭菌属的细菌被以下噬菌体感染:CAK1、CA5、Ca7、CEβ、(syn=1C)、CEγ、Cld1、c- n71、c-203Tox-、DEβ、(syn=1D)、(syn=1Dtox )、HM3、KM1、KT、Ms、NA1、(syn=Na1tox )、 PA1350e、 PL73、PL78、PL81、P1、P50、P5771、P19402、1Ctox 、2Ctox-、2D、(syn=2Dtox )、 3C、(syn=3Ctox )、4C、(syn=4tox )、56、III-1、NN-梭菌(61)、NB1tox-α1、CA1、HMT、HM2、 PF1、P-23、P-46、Q-05、Q-06、Q-16、Q-21、Q-26、Q-40、Q-46、S111、SA02、WA01、WA03、W111、 W523、80、C、CA2、CA3、CPT1、CPT4、c1、c4、c5、HM7、H11/A1、H18/A1、H22/S23、H158/A1、K2/A1、 K21/S23、ML、NA2tox-、Pf2、Pf3、Pf4、S9/S3、S41/A1、S44/S23、α2、41、112/S23、214/S23、 233/A1、234/S23、235/S23、II-1、II-2、II-3、NN-梭菌(12)、CA1、F1、K、S2、1、5和NN-梭菌 (8)。 棒状杆菌属的细菌被以下噬菌体感染:CGI1(缺陷型)、A、A2、A3、A110、A128、A133、 A137、A139、A155、A182、B、BF、B17、B18、B51、B271、B275、B276、B277、B279、B282、C、cap1、 CC1、CG1、CG2、CG33、CL31、Cog、(syn=CG5)、D、E、F、H、H-1、hq1、hq2、I1/H33、I1/H33、J、K、K、 (syn=Ktox-)、L、L、(syn=Ltox )、M、MC-1、MC-2、MC-3、MC-4、MLMa、N、O、ov1、ov2、ov3、P、P、 P、RP6、RS29、S、T、U、UB1、ub2、UH1、UH3、uh3、uh5、uh6、β、(syn=βtox )、βlaν64、βvir、γ、 (syn=γtox-)、γ19、δ、(syn=δtox )、ρ、(syn=ρtox-)、 ω、1Α、1/1180、2、2/ 1180、5/1180、5ad/9717、7/4465、8/4465、8ad/10269、10/9253、13/9253、15/3148、21/9253、 28、29、55、2747、2893、4498和5848。 肠球菌属的细菌被以下噬菌体感染:DF78、F1、F2、1、2、4、14、41、867、D1、SB24、 2BV、182、225、C2、C2F、E3、E62、DS96、H24、M35、P3、P9、SB101、S2、2BII、5、182a、705、873、 881、940、1051、1057、21096C、NN-肠球菌(1)、PE1、P1、F3、F4、VD13、1、200、235和341。 丹毒丝菌属的细菌被以下噬菌体感染:NN-丹毒丝菌(1)。 埃希氏菌属的细菌被以下噬菌体感染:BW73、B278、D6、D108、E、E1、E24、E41、FI-2、 FI-4、FI-5、HI8A、HI8B、i、MM、Mu、(syn=mu)、(syn=Mu1)、(syn=Mu-1)、(syn=MU-1)、(syn =MuI)、(syn=mu)、O25、PhI-5、Pk、PSP3、P1、P1D、P2、P4(缺陷型)、S1、 (缺陷 型)、 ψ(缺陷型)、7Α、 15(缺陷型)、18、28-1、186、299、NN-埃希氏菌 20 CN 111601897 A 说 明 书 18/43 页 (2)、ΑΒ48、CM、C4、C16、DD-VI、(syn=Dd-Vi)、(syn=DDVI)、(syn=DDVi)、E4、E7、E28、 F11、F13、H、H1、H3、H8、K3、M、N、ND-2、ND-3、ND4、ND-5、ND6、ND-7、Ox-1、(syn=OX1)、(syn= 11F)、Ox-2、(syn=Ox2)、(syn=OX2)、Ox-3、Ox-4、Ox-S、(syn=OX5)、Ox-6、(syn=66F)、 O111、Phl-1、RB42、RB43、RB49、RB69、S、Sal-1、Sal-2、Sal-3、Sal-4、 Sal-5、Sal-6、TC23、TC45、TuII*-6、(syn=TuII*)、TuII*-24、TuII*46、TuII*-60、T2、(syn =gamma)、(syn=γ)、(syn=PC)、(syn=P.C.)、(syn=T-2)、(syn=T2)、(syn=P4)、T4、 (syn=T-4)、(syn=T4)、T6、T35、α1、1、1A、3、(syn=Ac3)、3A、3T 、(syn=3)、(syn=M1)、 9266Q、CFO103、HK620、J、K、K1F、m59、no.A、no.E、no.3、no.9、N4、sd、(syn= Sd)、(syn=SD)、(syn=Sd)、(syn=sd)、(syn=SD)、(syn=CD)、T3、(syn=T-3)、(syn= T3)、T7、(syn=T-7)、(syn=T7)、WPK、W31、ΔΗ、 Φ04-CF、Φ 05、Φ06、Φ07、 Ω8、1、3、7、8、26、27、 28-2、29、30、31、32、38、39、42、933W、NN-埃希氏菌(1)、Esc-7-11、AC30、CVX-5、C1、DDUP、 EC1、EC2、E21、E29、F1、F26S、F27S、Hi、HK022、HK97、(syn=ΦHK97)、HK139、HK253、HK256、 K7、ND-1、no.D、PA-2、q、S2、T1、(syn=α)、(syn=P28)、(syn=T-1)、(syn=T1)、T3C、T5、 (syn=T-5)、(syn=T5)、UC-1、w、β4、γ2、λ、(syn=lambda)、(syn=Φλ)、ΦD326、 Φ06、 Φ7、Φ10、 χ、(syn=χ1)、 2、4、4Α、6、8Α、102、150、168、174、3000、 AC6、AC7、AC28、AC43、AC50、AC57、AC81、AC95、ΗK243、K10、ZG/3A、5、5Α、21EL、H19-f和 933H。 梭杆菌属的细菌被以下噬菌体感染:NN-梭杆菌(2)、fv83-554/3、fv88-531/2、 227、fv2377、fv2527和fv8501。 嗜血杆菌属的细菌被以下噬菌体感染:HP1(嗜血杆菌噬菌体HP1)、S2和N3。 螺杆菌属的细菌被以下噬菌体感染:HP1(幽门螺杆菌噬菌体HP1)和NN-螺杆菌 (1)。 克雷伯氏菌属的细菌被以下噬菌体感染:AIO-2、Kl4B、Kl6B、Kl9、(syn=Kl9)、 Kl14、Kl15、Kl21、Kl28、Kl29、Kl32、Kl33、Kl35、Kl106B、Kl171B、Kl181B、Kl832B、AIO-1、AO- 1、AO-2、AO-3、FC3-10、K、Kl1、(syn=Kl1)、Kl2、(syn=Kl2)、Kl3、(syn=Kl3)、(syn=Kl70/ 11)、Kl4、(syn=Kl4)、Kl5、(syn=Kl5)、Kl6、(syn=Kl6)、Kl7、(syn=Kl7)、Kl8、(syn= Kl8)、Kl19、(syn=Kl9)、Kl27、(syn=K127)、Kl31、(syn=Kl31)、Kl35、Kl171B、II、VI、IX、 CI-1、Kl4B、Kl8、Kl11、Kl12、Kl13、Kl16、Kl17、Kl18、Kl20、Kl22、Kl23、Kl24、Kl26、Kl30、 Kl34、Kl106B、Kl165B、Kl328B、KLXI、K328、P5046、11、380、III、IV、VII、VIII、FC3-11、Kl2B、 (syn=Kl2B)、Kl25、(syn=Kl25)、Kl42B、(syn=Kl42)、(syn=Kl42B)、Kl181B、(syn= Kl181)、(syn=Kl181B)、Kl765/1、(syn=Kl765/1)、Kl842B、(syn=Kl832B)、Kl937B、(syn =Kl937B)、L1、φ28、7、231、483、490、632和864/100。 钩端螺旋体属的细菌被以下噬菌体感染:LE1、LE3、LE4和NN-钩端螺旋体(1)。 李斯特菌属的细菌被以下噬菌体感染:A511、O1761、4211、4286、(syn=BO54)、 A005、A006、A020、A500、A502、A511、A118、A620、A640、B012、B021、B024、B025、B035、B051、 B053、B054、B055、B056、B101、B110、B545、B604、B653、C707、D441、HSO47、H1OG、H8/73、H19、 H21、H43、H46、H107、H108、H10、H163/84、H1312、H340、H387、H391/73、H684/74、H924A、PSA、 21 CN 111601897 A 说 明 书 19/43 页 U153、 (syn=P35)、00241、00611、02971A、02971C、5/476、5/911、5/939、5/11302、5/ 11605、5/11704、184、575、633、699/694、744、900、1090、1317、1444、1652、1806、1807、1921/ 959、1921/11367、1921/11500、1921/11566、1921/12460、1921/12582、1967、2389、2425、 2671、2685、3274、3550、3551、3552、4276、4277、4292、4477、5337、5348/11363、5348/11646、 5348/12430、5348/12434、10072、11355C、11711A、12029、12981、13441、90666、90816、 93253、907515、910716和NN-李斯特菌(15)。 摩根氏菌属的细菌被以下噬菌体感染:47。 分枝杆菌属的细菌被以下噬菌体感染:I3、AG1、AL1、ATCC 11759、A2、B.C3、BG2、 BK1、BK5、乳酪分枝杆菌、B-1、B5、B7、B30、B35、Clark、C1、C2、DNAIII、DSP1、D4、D29、GS4E、 (syn=GS4E)、GS7、(syn=GS-7)、(syn=GS7)、IPα、lacticola、Legendre、Leo、L5、(syn=Φ L-5)、MC-1、MC-3、MC-4、minetti、MTPH11、Mx4、MyF3P/59a、phlei、(syn=phlei 1)、phlei 4、Polonus II、rabinovitschi、smegmatis、TM4、TM9、TM10、TM120、Y7、Y10、 1B、1F、1H、 1/1、67、106、1430、B1、(syn=Bol)、B24、D、D29、F-K、F-S、HP、Polonus I、Roy、R1、(syn=R1- Myb)、(syn=R1)、11、31、40、50、103a、103b、128、3111-D、3215-D和NN-分枝杆菌(1)。 奈瑟氏菌属的细菌被以下噬菌体感染:I组、II组和NP1。 诺卡氏菌属的细菌被以下噬菌体感染:P8、NJ-L、NS-8、N5和NN-诺卡氏菌(1)。 变形杆菌属的细菌被以下噬菌体感染:Pm5、13vir、2/44、4/545、6/1004、13/807、 20/826、57、67b、78、107/69、121、9/0、22/608、30/680、Pm1、Pm3、Pm4、Pm6、Pm7、Pm9、Pm10、 Pm11、Pv2、π1、 7/549、9B/2、10A/31、12/55、14、15、16/789、17/971、19A/653、23/532、 25/909、26/219、27/953、32A/909、33/971、34/13、65、5006M、7480b、VI、13/3a、Clichy 12、π 2600、 1/1004、5/742、9、12、14、22、24/860、2600/D52、Pm8和24/2514。 普罗维登斯菌属的细菌被以下噬菌体感染:PL25、PL26、PL37、9211/9295、9213/ 9211b、9248、7/R49、74761322、7478/325、7479、7480、9000/9402和9213/9211a。 假单胞菌属的细菌被以下噬菌体感染:Pf1、(syn=Pf-1)、Pf2、Pf3、PP7、PRR1、7s、 NN-假单胞菌(1)、AI-1、M-2、B17、B89、CB3、Col 2、Col 11、Col 18、Col 21、C154、C163、 C167、C2121、E79、F8、ga、gb、H22、K1、M4、N2、Nu、PB-1、(syn=PB1)、pf16、PMN17、PP1、PP8、 Psa1、PsP1、PsP2、PsP3、PsP4、PsP5、PS3、PS17、PTB80、PX4、PX7、PYO1、PYO2、PYO5、PYO6、 PYO9、PYO10、PYO13、PYO14、PYO16、PYO18、PYO19、PYO20、PYO29、PYO32、PYO33、PYO35、PYO36、 PYO37、PYO38、PYO39、PYO41、PYO42、PYO45、PYO47、PYO48、PYO64、PYO69、PYO103、PlK、SLP1、 SL2、S2、UNL-1、wy、Ya1、Ya4、Ya11、 (syn=ΦKZ)、 Φmu78、 1/72、2/79、3、3/DO、4/237、5/406、6C、6/6660、7、7v、7/ 184、8/280、9/95、10/502、11/DE、12/100、12S、16、21、24、25F、27、31、44、68、71、95、109、 188、337、352、1214、NN-假单胞菌(23)、A856、B26、CI-1、CI-2、C5、D、gh-1、F116、HF、H90、K5、 K6、K104、K109、K166、K267、N4、N5、O6N-25P、PE69、Pf、PPN25、PPN35、PPN89、PPN91、PP2、PP3、 PP4、PP6、PP7、PP8、PP56、PP87、PP114、PP206、PP207、PP306、PP651、Psp231a、Pssy401、 Pssy9220、ps1、PTB2、PTB20、PTB42、PX1、PX3、PX10、PX12、PX14、PYO70、PYO71、R、SH6、SH133、 tf、Ya5、Ya7、 ΦKf77、 ΦmnF82、 1、2、2、3、4、5、6、7、7、8、9、 10、11、12、12B、13、14、15、14、15、16、17、18、19、20、20、21、21、22、23、23、24、25、31、53、73、 119x、145、147、170、267、284、308、525、NN-假单胞菌(5)、af、A7、B3、B33、B39、BI-1、C22、D3、 22 CN 111601897 A 说 明 书 20/43 页 D37、D40、D62、D3112、F7、F10、g、gd、ge、gf、Hw12、Jb19、KF1、L°、OXN-32P、O6N-52P、PCH-1、 PC13-1、PC35-1、PH2、PH51、PH93、PH132、PMW、PM13、PM57、PM61、PM62、PM63、PM69、PM105、 PM113、PM681、PM682、PO4、PP1、PP4、PP5、PP64、PP65、PP66、PP71、PP86、PP88、PP92、PP401、 PP711、PP891、Pssy41、Pssy42、Pssy403、Pssy404、Pssy420、Pssy923、PS4、PS-10、Pz、SD1、 SL1、SL3、SL5、SM、 Φc11、 2、2F、5、7m、 11、13、13/441、14、20、24、40、45、49、61、73、148、160、198、218、222、236、242、246、249、258、 269、295、297、309、318、342、350、351、357-1、400-1、NN-假单胞菌(6)、G10、M6、M6a、L1、PB2、 Pssy15、Pssy4210、Pssy4220、PYO12、PYO34、PYO49、PYO50、PYO51、PY052、PYO53、PYO57、 PYO59、PYO200、PX2、PX5、SL4、 和1214。 立克次氏体属的细菌被以下噬菌体感染:NN-立克次氏体(1)。 沙门氏菌属的细菌被以下噬菌体感染:b、Beccles、CT、d、Dundee、f、Fels 2、GI、 GIII、GVI、GVIII、k、K、i、j、L、O1、(syn=O-1)、(syn=O1)、(syn=O-I)、(syn=7)、O2、O3、 P3、P9a、P10、Sab3、Sab5、San15、San17、SI、Taunton、ViI、(syn=Vil)、9、NN-沙门氏菌(1)、 N-1、N-5、N-10、N-17、N-22、11、12、16-19、20.2、36、449C/C178、966A/C259、a、B.A.O.R.、e、 G4、GIII、L、LP7、M、MG40、N-18、PSA68、P4、P9c、P22、(syn=P22)、(syn=PLT22)、(syn= PLT22)、P22a1、P22-4、P22-7、P22-11、SNT-1、SNT-2、SP6、ViIII、ViIV、ViV、ViVI、ViVII、 Worksop、ε15、ε34、1、37、1(40)、(syn=φ1[40])、1、422、2、2.5、3b、4、5、6、14(18)、8、14(6, 7)、10、27、28B、30、31、32、33、34、36、37、39、1412、SNT-3,7-11、40.3、c、C236、C557、C625、 C966N、g、GV、G5、G173、h、IRA、Jersey、M78、P22-1、P22-3、P22-12、Sab1、Sab2、Sab2、Sab4、 San1、San2、San3、San4、San6、San7、San8、San9、San13、San14、San16、San18、San19、San20、 San21、San22、San23、San24、San25、San26、SasL1、SasL2、SasL3、SasL4、SasL5、S1BL、SII、 Vill、φ1、1、2、3a、3aI、1010、NN-沙门氏菌(1)、N-4、SasL6和27。 沙雷氏菌属的细菌被以下噬菌体感染:A2P、PS20、SMB3、SMP、SMP5、SM2、V40、V56、 κ、DCP-3、(DCP-6、3M、10/1a、20A、34CC、34H、38T、345G、345P、501B、SMB2、SMP2、BC、BT、CW2、 CW3、CW4、CW5、L1232、L2232、L34、L.228、SLP、SMPA、V.43、σ、 ΦCP6-1、ΦCP6-2、ΦCΡ 6-5、3T、5、8、9F、10/1、20E、32/6、34B、34CT、34P、37、41、56、56D、56P、60P、61/6、74/6、76/4、 101/8900、226、227、228、229F、286、289、290F、512、764a、2847/10、2847/10a、L.359和SMB1。 志贺氏菌属的细菌被以下噬菌体感染:Fsa、(syn=a)、FSD2d、(syn=D2d)、(syn= W2d)、FSD2E、(syn=W2e)、fv、F6、f7.8、H-Sh、PE5、P90、SfII、Sh、SHIII、SHIV、(syn=HIV)、 SHVI、(syn=HVI)、SHVIII、(syn=HVIII)、SKγ66、(syn=gamma 66)、(syn=γ66)、(syn= γ66b)、SKIII、(syn=SIIIb)、(syn=III)、SKIV、(syn=SIVa)、(syn=lV)、SKIVa、(syn= SIVAn)、(syn=IVA)、SKVI、(syn=KVI)、(syn=SVI)、(syn=VI)、SKVIII、(syn=SVIII)、 (syn=VIII)、SKVIIIA、(syn=SVIIIA)、(syn=VIIIA)、STVI、STIX、STXI、STXII、S66、W2、 (syn=D2c)、(syn=D20)、 ΦIV1、3-SO-R、8368-SO-R、F7、(syn=FS7)、(syn=K29)、F10、 (syn=FS10)、(syn=K31)、I1、(syn=alfa)、(syn=FSα)、(syn=K18)、(syn=α)、I2、(syn =a)、(syn=K19)、SG35、(syn=G35)、(syn=SO-35/G)、SG55、(syn=SO-55/G)、SG3201、 (syn=SO-3201/G)、SHII、(syn=HII)、SHV、(syn=SHV)、SHX、SHX、SKII、(syn=K2)、(syn= KII)、(syn=SII)、(syn=SsII)、(syn=II)、SKIV、(syn=SIVb)、(syn=SsIV)、(syn=IV)、 SKIVa、(syn=SIVab)、(syn=SsIVa)、(syn=lVa)、SKV、(syn=K4)、(syn=KV)、(syn=SV)、 23 CN 111601897 A 说 明 书 21/43 页 (syn=SsV)、(syn=V)、SKX、(syn=K9)、(syn=KX)、(syn=SX)、(syn=SsX)、(syn=X)、 STV、(syn=T35)、(syn=35-50-R)、STVIII、(syn=T8345)、(syn=8345-SO-S-R)、W1、(syn =D8)、(syn=FSD8)、W2a、(syn=D2A)、(syn=FS2a)、DD-2、Sf6、FS1、(syn=F1)、SF6、(syn =F6)、SG42、(syn=SO-42/G)、SG3203、(syn=SO-3203/G)、SKF12、(syn=SsF12)、(syn= F12)、(syn=F12)、STII、(syn=1881-SO-R)、γ66、(syn=gamma 66a)、(syn=Ssγ66)、 B11、DDVII、(syn=DD7)、FSD2b、(syn=W2B)、FS2、(syn=F2)、(syn=F2)、FS4、(syn=F4)、 (syn=F4)、FS5、(syn=F5)、(syn=F5)、FS9、(syn=F9)、(syn=F9)、F11、P2-SO-S、SG36、 (syn=SO-36/G)、(syn=G36)、SG3204、(syn=SO-3204/G)、SG3244、(syn=SO-3244/G)、 SHI、(syn=HI)、SHVII、(syn=HVII)、SHIX、(syn=HIX)、SHXI、SHXII、(syn=HXII)、SKI、 KI、(syn=SI)、(syn=SsI)、SKVII、(syn=KVII)、(syn=SVII)、(syn=SsVII)、SKIX、(syn =KIX)、(syn=SIX)、(syn=SsIX)、SKXII、(syn=KXII)、(syn=SVII)、(syn=SsXII)、STI、 SIII、STIII、STIV、STVII、S70、S206、U2-SO—S、3210-SO-S、3859-SO-S、4020-SO-S、 SHIII、(syn=HIII)、SHXI、(syn=HXI)和SXI、(syn= KXI)、(syn=SXI)、(syn=SsXI)、(syn=XI)。 葡萄球菌属的细菌被以下噬菌体感染:A、EW、K、Ph5、Ph9、Ph10、Ph13、P1、P2、P3、 P4、P8、P9、P10、RG、SB-1、(syn=Sb-1)、S3K、Twort、 06、40、58、119、130、131、 200、1623、STC1、(syn=stc1)、STC2、(syn=stc2)、44AHJD、68、AC1、AC2、A6"C"、A9"C"、 b581、CA-1、CA-2、CA-3、CA-4、CA-5、D11、L39x35、L54a、M42、N1、N2、N3、N4、N5、N7、N8、N10、 N11、N12、N13、N14、N16、Ph6、Ph12、Ph14、UC-18、U4、U15、S1、S2、S3、S4、S5、X2、Z1、 ω、11、 (syn=P11-M15)、15、28、28A、29、31、31B、37、42D、(syn=P42D)、44A、48、 51、52、52A、(syn=P52A)、52B、53、55、69、71、(syn=P71)、71A、72、75、76、77、79、80、80a、 82、82A、83A、84、85、86、88、88A、89、90、92、95、96、102、107、108、111、129-26、130、130A、 155、157、157A、165、187、275、275A、275B、356、456、459、471、471A、489、581、676、898、1139、 1154A、1259、1314、1380、1405、1563、2148、2638A、2638B、2638C、2731、2792A、2792B、2818、 2835、2848A、3619、5841、12100、AC3、A8、A10、A13、b594n、D、M12、N9、N15、P52、P87、S1、S6、 Z4、 3A、3B、3C、6、7、16、21、42B、42C、42E、44、47、47A、47C、51、54、54x1、70、73、75、78、 81、82、88、93、94、101、105、110、115、129/16、174、594n、1363/14、2460和NN-葡萄球菌(1)。 链球菌属的细菌被以下噬菌体感染:EJ-1、NN-链球菌(1)、a、CI、FLOThs、H39、Cp- 1、Cp-5、Cp-7、Cp-9、Cp-10、AT298、A5、a10/J1、a10/J2、a10/J5、a10/J9、A25、BT11、b6、CA1、 c20-1、c20-2、DP-1、Dp-4、DT1、ET42、e10、FA101、FEThs、FK、FKK101、FKL10、FKP74、FK11、 FLOThs、FY101、f1、F10、F20140/76、g、GT-234、HB3、(syn=HB-3)、HB-623、HB-746、M102、 O1205、 PST、P0、P1、P2、P3、P5、P6、P8、P9、P9、P12、P13、P14、P49、P50、P51、P52、P53、 P54、P55、P56、P57、P58、P59、P64、P67、P69、P71、P73、P75、P76、P77、P82、P83、P88、sc、sch、 sf、Sfi11、(syn=SFi11)、 sfi19、(syn=SFi19)、 Sfi21、(syn=SFi21)、 STG、STX、st2、ST2、 ST4、S3、 s265、Φ17、 Φ57、 Φ7201、ω1、ω2、ω3、ω4、ω5、ω6、ω8、ω10、1、6、9、 24 CN 111601897 A 说 明 书 22/43 页 10F、12/12、14、17SR、19S、24、50/33、50/34、55/14、55/15、70/35、70/36、71/ST15、71/45、 71/46、74F、79137、79/38、80/J4、80/J9、80/ST16、80/15、80/47、80/48、101、103/39、103/ 40、121/41、121/42、123/43、123/44、124/44、337/ST17和NN-链球菌(34)。 密螺旋体属的细菌被以下噬菌体感染:NN-密螺旋体(1)。 弧菌属的细菌被以下噬菌体感染:CΤXΦ、fs、(syn=s1)、fs2、1vpfs、Vf12、Vf33、 ΥΡΙΦ、VSK、v6、493、CP-T1、ET25、kappa、K139、LaboI,)XN-69P、OXN-86、O6N-21P、PB-1、 P147、rp-1、SE3、VA-1、(syn=VcA-1)、VcA-2、VcA-1、VP1、VP2、VP4、VP7、VP8、VP9、VP10、 VP17、VP18、VP19、X29、(syn=29d’Hérelle)、1、ΦHAWI-1、ΦHAWI-2、ΦHAWI-3、ΦHAWI-4、 ΦHAWI-5、ΦHAWI-6、ΦHAWI-7、ΦHAWI-8、ΦHAWI-9、ΦHAWI-10、ΦHC1-1、ΦHC1-2、ΦHC1- 3、ΦHC1-4、ΦHC2-1、ΦHC2-2、ΦHC2-3、ΦHC2-4、ΦHC3-1、ΦHC3-2、ΦHC3-3、ΦHD1S-1、Φ HD2S-1、ΦHD2S-2、ΦHD2S-3、ΦHD2S-4、ΦHD2S-5、ΦHDO-1、ΦHDO-2、ΦHDO-3、ΦHDO-4、Φ HDO-5、ΦHDO-6、ΦKL-33、ΦKL-34、ΦKL-35、ΦKL-36、ΦKW1H-2、ΦKWH-3、ΦKWH-4、Φ MARQ-1、ΦMARQ-2、ΦMARQ-3、ΦMOAT-1、ΦO139、ΦPEL1A-1、ΦPEL1A-2、ΦPEL8A-1、Φ PEL8A-2、ΦPEL8A-3、ΦPEL8C-1、ΦPEL8C-2、ΦPEL13A-1、ΦPEL13B-1、ΦPEL13B3-2、Φ PEL13B-3、ΦPEL13B-4、ΦPEL13B-5、ΦPEL13B-6、ΦPEL13B-7、ΦPEL13B-8、ΦPEL13B-9、Φ PEL13B-10、 Φ16、 1-11、5、13、14、16、24、32、493、6214、7050、7227、 II、(syn=组11)、 V、VIII、NN-弧菌(13)、KVP20、KVP40、nt-1、O6N-22P、P68、e1、e2、 e3、e4、e5、FK、G、J、K、nt-6、N1、N2、N3、N4、N5、O6N-34P、OXN-72P、OXN-85P、OXN-100P、P、Ph- 1、PL163/10、Q、S、T、 1-9、37、51、57、70A-8、72A-4、72A-10、110A-4、333、4996、I、(syn= 组I)、ΙII、(syn=组III)、VI、(syn=A-Saratov)、VII、IX、X、NN-弧菌(6)、pA1、7、7-8、70A- 2、71A-6、72A-5、72A-8、108A-10、109A-6、109A-8、110A-1、110A-5、110A-7、hv-1、OXN-52P、 P13、P38、P53、P65、P108、P111、TP1、VP3、VP6、VPI2、VP13、70A-3、70A-4、70A-10、72A-1、 108A-3、109-B1、110A-2、149、 IV、(syn=组IV)、NN-弧菌(22)、VP5、VP11、VP15、 VP16、α1、α2、α3a、3b、353B和NN-弧菌(7)。 耶尔森氏菌属的细菌被以下噬菌体感染:H、H-1、H-2、H-3、H-4、Lucas 110、Lucas 303、Lucas 404、YerA3、YerA7、YerA20、YerA41、3/M64-76、5/G394-76、6/C753-76、8/C239- 76、9/F18167、1701、1710、PST、1/F2852-76、D’Hérelle、EV、H、Kotljarova、PTB、R、Y、 YerA41、 3、4/C1324-76、7/F783-76、903、1/M6176和Yer2AT。 在另一实施方案中,使用至少2种、至少3种、至少4种、至少5种、至少6种、至少7种、 至少8种、至少9种或更多种上述噬菌体的组合来实施所述方法。本领域技术人员认识到,转 化效率可以操纵,例如,增强或抑制,这取决于所用噬菌体的特定组合。 特别地,细菌物种(和相应的宿主特异性噬菌体)包括嗜水气单胞菌(Aeromonas hydrophila) (PM2)、炭疽芽孢杆菌(Bacillus anthracis) (Gamma)、枯草芽孢杆菌 (Bacillus subtilus) (SPP1)、百日咳博德特氏菌(Bordetella pertussis) (参见 Pereversev等Zh Mikrobiol 5:54-57 ,1981)、伯氏疏螺旋体(Borrelia burgdorferi)( 参见Eggers等,JBacteriol 183:4771-4778 ,2001)、流产布鲁氏菌(Brucella abortus)(TB;212;371)、空肠弯曲杆菌(Campylobacter jejuni) 产气荚膜梭菌 (Clostridium perfringes) 粪肠球菌(Enterococcus faecalis) 屎肠球菌 25 CN 111601897 A 说 明 书 23/43 页 (Enterococcus faecium)(ENB6)、大肠杆菌(Escherichia coli)(P1;T1;T3、T4、T5;T7、 KH1、 lambda; mu)、肺炎克雷伯菌(Klebsiella pneumoniae)(60;92)、单核细胞增 生性李斯特菌(Listeria monocytogenes)(A511、A118;243;H387;2389;2671;2685;4211)、 麻风分枝杆菌(Mycobacterium leprae) (分枝杆菌噬菌体,L5)、结核分枝杆菌 (Mycobacterium tuberculosis)(LG;DSGA)、铜绿假单胞菌(Pseudomonas aeruginosa) (E79,G101;B3;pp.7)、鸭沙门氏菌(Salmonella anatum)(E5)、病牛沙门氏菌(Salmonella bovismorbificans)(98)、猪霍乱沙门氏菌(Salmonella choleraesuis)(102)、肠炎沙门氏 菌(Salmonella enteritidis)(L;P22;102;FO;IRA; )、纽因吞沙门氏菌(Salmonella Newington)(E34)、肖特苗勒氏沙门氏菌(Salmonella schottmulleri)(31;102;F0;14)、伤 寒沙门氏菌(Salmonella typhi)(163;175;ViI;ViVI;8;23;25;46;175;F0)、粘质沙雷氏菌 (Serratia marcescens)(S24VA)、志贺氏痢疾杆菌(Shigella dysenteriae)( P2;2; 37)、福氏志贺氏菌(Shigella flexneri) (Sf6)、金黄色葡萄球菌(Staphylococcus aureus)(K;P1;P14;UC18;15;17;29;42D;47;52;53;79;80;81;83A;92;Twort, )、肺炎链 球菌(Streptococcus pneumoniae)(Dp-1;Cp-1;HB-3;EJ-1;MM1;VO1)、化脓性链球菌 (Streptococcus pyogenes)( 1A;1B;T12;12/12;113;120;124;P58;H4489a)、霍乱弧 菌(Vibrio cholerae)(138;145;149;163)和鼠疫耶尔森氏菌(Yersinia pestis)(A1122; R;Y;P1)。另外的信息在美国专利公开No.2009-0155768中提供。 特别地,表1-3提供了特定宿主特异性噬菌体及其特异性的宿主的代表性实例,包 括它们介导其作用的受体。还参见Bertozzi等,FEMS Microbiology Letters,363,1-11 , 2016。 表1.格兰氏阳性细菌的细胞壁中的受体。宿主名称按字母顺序排序。 26 CN 111601897 A 说 明 书 24/43 页 表2.格兰氏阴性细菌的细胞壁中的受体。宿主名称按字母顺序排序。 27 CN 111601897 A 说 明 书 25/43 页 表3.细胞壁以外的细胞复合物中的受体。宿主名称按字母顺序排序。 28 CN 111601897 A 说 明 书 26/43 页 D.噬菌体包装位点 噬菌体包装位点是噬菌体基因组上用于基因组包装到病毒颗粒中的特定DNA序 列。质粒工程化以含有噬菌体包装位点,使得质粒包装到转导颗粒中。宿主特异性噬菌体 (及其包装位点)包括但不限于SPP1(SPP1 pac位点)、P1(P1 pac位点)、T1(T1 pac位点)、T7 (T7多联体接合)、lambda(cos位点)、mu(mu pac位点)、P22(P22 pac位点)、 ( pac位 点)、Sf6(Sf6 pac位点)、149(149pac位点)和A1122(A1122-多联体接合)。对于大多数噬菌 体,包装位点称为pac位点。在一些情况下,包装位点被称为多联体接合(例如,T7多联体接 合)。在每种情况下,包装位点不同于噬菌体基因组中天然存在的相邻序列。 对于一些噬菌体,包装位点可能是未知的。在这些情况下,pac位点可以通过利用 包含功能性噬菌体pac位点的质粒被包装的特性来确定。例如,通过将λ噬菌体基因组DNA的 小的限制性片段并入质粒中来确定噬菌体λ包装所必需的DNA序列(Hohn等,PNAS USA 80: 7456-7460,1983)。在引入体内包装菌株后,定量评估包装/转导的效率。使用类似的策略, 确定了许多噬菌体的pac位点:λ(Miwa等,Gene 20:267-279 ,1982);Mu(Croenen等, Virology 144:520-522,1985);丝状噬菌体,包括f1、fd、M13和Ike(Russel等,J Virol., 63:3284-3295,1989);P22(Petri等,Gene 88:47-55,1990;Wu等,Mol.Microbiol 45:1631- 1646,2002);T7(Chung等,J Mol Biol 216:927-938,1990)和T3(Hashimoto等,Virology 187:788-795,1992)。 29 CN 111601897 A 说 明 书 27/43 页 该方法的实施方案包括噬菌体包装序列及其功能片段。这些核酸实施方案可以 是,例如,长度至少9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、 29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、 54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、 79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、125、150、 175、200、250、300、350、400、450、500、550、600、650、700、750、800、850和900个核苷酸,只要 核苷酸片段可以介导质粒DNA包装到噬菌体衣壳中(如通过其介导包装,从而产生功能性转 导颗粒的能力判断)。将包含噬菌体包装位点或其片段的核酸并入质粒中。 E.标志物基因构建体 基因技术广泛用于监测细胞基因表达(Naylor等,Biochem Pharm 58:749-757 , 1990)。优选地,标志物分子是编码可检测产物(例如蛋白质或酶)的基因。特别优选地,标志 物是不被噬菌体或细菌天然表达的分子。例如,标志物可以是异源真核蛋白、不同细菌物种 的蛋白质或病毒蛋白质。 在一个实施方案中,标志物是抗原、酶、抗体或其片段以及适体。如本文所用,术语 “抗原”是指包含一个或多个表位(线性,构象或两者)的分子,该表位促进与结合配体(例 如,抗体)的特异性结合。术语“抗原”也可以指抗体(例如抗独特型抗体或其片段),以及指 可以模拟抗原或抗原决定簇(表位)的合成肽模拟表位。术语“抗原”还可以指在体内表达抗 原或抗原决定簇的寡核苷酸或多核苷酸。如本文所用,术语“表位”通常是指抗原上被抗体 或T细胞受体识别的位点。它可以是衍生自蛋白质抗原或作为蛋白质抗原的部分的短肽。然 而,该术语还旨在包括具有糖肽和碳水化合物表位的肽。几个不同的表位可以由单个抗原 分子携带。术语“表位”还包括修饰的氨基酸序列或碳水化合物,其刺激识别整个生物体的 反应。 在另一实施方案中,标志物是抗体。术语“抗体”包括整个抗体分子以及其抗原结 合片段。抗体包括IgG、IgA、IgM、IgE、IgD以及抗体变体,例如单链抗体(scFv)。合适的抗体 片段包含抗原结合位点,且因此包括但不限于Fv、Fab和F(ab)2片段。抗体可以是多克隆抗 体或单克隆抗体。在优选的实施方案中,抗体是单克隆抗体。还包括嵌合或合成抗体。在特 别优选的实施方案中,抗体是与目标抗原特异性结合的scFv。术语“特异性结合”是指抗体 与特定靶表位的结合水平(“信号”)高于其他非靶标(“噪音”)。当用于检测的信噪比为至少 0.6倍、0.7倍、0.8倍、0.9倍、1倍(100%增加)、1.5倍、3倍、5倍、10倍、20倍、50倍、70倍、100 倍或更多时,实现特异性检测。用于制备和表征抗体的方法是本领域熟知的(参见,例如, Lane等,“Antibodies:A Laboratory Manual”Cold Spring Harbor Laboratory,1988)。 在又一个实施方案中,标志物是适体。本文使用的术语“适体”是指对于特定材料 具有高特异性和亲和力的单链DNA(ssDNA)或RNA。由于制造昂贵的抗体试剂的限制,免疫检 测方法是昂贵且缓慢的。另一方面,由于适体使用相对简单的方法合成,并且细胞、蛋白质 和小有机材料可以是靶材料,因此可以开发使用适体的新的检测方法,并且其特异性和稳 定性与开发的抗体相当。鉴于这些优点,DNA适体可用于蛋白质标志物的特异性检测。一般 公认的是,可以通过体外进化方法合成和/或鉴定对于其与多肽的结合是特异性的适体。用 于制备和表征适体的方法,包括通过体外进化方法,是本领域熟知的(例如美国专利号7, 939,313)。 30 CN 111601897 A 说 明 书 28/43 页 本方法部分地包括确定样品中至少一种标志物或指示物的存在(或不存在)或者 水平(例如,浓度)或活性(例如,酶活性)。如本文所用,术语“标志物”或“指示物”是指编码 核酸(例如,mRNA)、肽或蛋白质的核苷酸序列,其允许确定或确认载体已被正确地转染或转 导且其序列正确表达。标志物可以是编码蛋白质的核苷酸序列或编码抗生素抗性的基因, 其用于选择携带载体的细胞。如本文所用,术语“检测至少一种标志物的存在”包括通过使 用本领域技术人员已知的任何定量或定性试验来确定每种目标标志物的存在。在某些情况 下,确定特定性状、变量、基因型和/或生物化学或血清学物质(例如,蛋白质或抗体)的存在 或不存在的定性试验适合于检测每种目标标志物。在某些其他情况中,确定DNA、RNA、蛋白 质、抗体或活性的存在或不存在的定量试验适合于检测每种目标标志物。如本文所用,术语 “检测至少一种标志物的水平”包括通过使用本领域技术人员已知的任何直接或间接定量 试验来确定每种目标标志物的水平。在某些情况下,测定例如DNA、RNA、蛋白质、抗体或活性 的相对或绝对量的定量试验适合于检测每种目标标记物的水平。本领域技术人员将理解, 可用于检测标志物水平的任何试验也可用于检测标志物的存在或不存在。 在一些实施方案中,标志物是报告分子,其可以例如通过其发光特性或其进行酶 促反应的能力表示其存在。在另一实施方案中,标志物与报告分子结合以表示标志物的水 平或活性。在后一种情况下,报告体可以是结合标志物蛋白的抗体或配体。 (i)报告体 在一个实施方案中,报告分子是称为报告基因的基因,其编码可检测蛋白质的表 达。常用的报告基因包括氯霉素乙酰转移酶(CAT)、13-半乳糖苷酶、荧光素酶、碱性磷酸酶 和绿色荧光蛋白(GFP)。通常,报告基因具有低背景活性和基因表达后灵敏的信号检测的优 点。例如,荧光素酶和GFP作为基因表达的非侵入性标志物的开发,结合使用敏感电荷耦合 器件(CCD)成像相机和荧光显微镜的检测简易性,允许关于甚至在单细胞水平的基因表达 的时间和空间信息。 荧光素酶基因及其作为报告基因的用途的综述提供了哈维氏弧菌(Vibrio harveyi)(登录号M10961和AAA88685)、哈维氏弧菌(登录号M10961和AAA88686)、哈维氏弧 菌(登录号M28815和AAA27531)、费氏弧菌(Vibrio fischeri)(登录号X06758和CAA29931)、 费氏弧菌(登录号X06797和CAA29932)、费氏弧菌(登录号AF170104(包括其变体))、发光杆 菌(Photorhabdus luminescens)(登录号M62917)、北斗萤火虫(Photinus pyralis) (M15077和AAA29795)、源氏萤(Luciola cruciate)(登录号M26194和AAA29135)、希氏弯喉 海萤(Vargula hilgendorfii)(登录号E02749、M25666和AAA30332)、维多利亚多管发光水 母(Aequorea victoria)(登录号M16103、AAA27719、M11394、AAA27716、M16104、AAA27717、 M16105、AAA27718、L29571、AAA27720和E02319);细脚刺虾(Oplophorus gracilorostris) (登录号AB030246、BAB13776、AB030245和BAB13775);米氏肾海鳃(Renilla muelleri)(登 录号AY015988和AAG54094);和海肾(Renilla reniformis)(登录号M63501和AAA29804)的 已知荧光素酶基因、cDNA、蛋白质和相应的GENBANK登录号的列表。参见Greer等, Luminescence 17:43-74,2002)。Greer还提供了大量可用于成像的构建体和载体(参见表 2,第48-52页)。这些载体适合在金黄色葡萄球菌、大肠杆菌和其他细菌中表达。已知的荧光 素酶有原核荧光素酶(Lux)和真核荧光素酶(Luc、Ruc及其调节蛋白),这两者通常用于活细 胞中荧光素酶表达的成像。 31 CN 111601897 A 说 明 书 29/43 页 在另一实施方案中,报告分子包括在细菌中表达的β-半乳糖苷酶报告基因 (Miller等,Experiments in Molecular Genetics,Cold Spring Harbor Laboratory, Cold Spring Harbor,N .Y .;Sambrook等,Molecular Cloning,Cold Spring Harbor Laboratory,Cold Spring Harbor,N.Y.)。通过在含有X-Gal(5-溴-4-氯-3-吲哚基-β-D-吡 喃半乳糖苷)的培养基上的蓝色着色检测由细菌菌落表达的β-半乳糖苷酶活性。氯霉素乙 酰转移酶(CAT)也适合用作细菌中的报告基因。CAT由细菌药物抗性基因编码(Kondo等,J Bacterid 88:1266-1276)。它通过在两个羟基中的一个或两个处使药物乙酰化来灭活氯霉 素。在典型的CAT试验中,将细胞提取物在含有14C-或3H-标记的氯霉素和正丁酰辅酶A的反 应混合物中孵育。CAT将辅因子的正丁酰基部分转移至氯霉素。反应产物用二甲苯萃取,且 正丁酰基氯霉素主要分配到二甲苯相中,而未修饰的氯霉素主要保留在水相中。使用闪烁 计数器测量分配到二甲苯相中的放射性标记的氯霉素。 由大肠杆菌的phoA编码的细菌碱性磷酸酶仅在其通过细胞膜转运到周质空间中 时才具有酶活性(Gibson等,Appl and Env Microbiol 68:928-932,2002)。该特性已被用 于将PhoA蛋白工程化为亚细胞定位的分子传感器。源自革兰氏阳性细菌粪肠球菌的另一种 细菌碱性磷酸酶(PhoZ)(Lee等,J Bacteriol 181:5790-5799,1999)已被开发为在革兰氏 阳性细菌中高活性的报告基因(Granok等.J Bacteriol 182:1529-1540 ,2000;Lee等,J Bacteriol 181:5790-5799,1999)。PhoZ的碱性磷酸酶活性,与PhoA的碱性磷酸酶活性一 样,依赖于其从细胞质的输出。在碱性磷酸酶试验中,碱性磷酸酶水解底物如4-硝基苯基磷 酸酯(4NPP)以产生色原体(例如4-硝基苯酚,4NP)。 报告基因允许更简单的操作程序(例如减少纯化或细胞裂解),它们可适用于大规 模、高通量的筛选测量,并且它们与细菌系统相容。报告基因可以是天然存在的基因或通过 遗传操作(如重组DNA技术或诱变)产生的基因。报告基因是含有编码区和任何相关表达序 列如启动子、翻译起始序列和调控序列的核酸区段。 (ii)细菌特异性启动子 报告基因通常与控制和指导RNA合成的启动子序列连接。本领域普通技术人员应 理解,启动子序列可选自由各种细菌物种表达的大量细菌基因。启动子的选择基于待检测 的目标细菌进行。对于在大肠杆菌中实现高水平基因表达的策略的综述,参见Makrides等, Microbiol Rev 60:512-538,1996。在大肠杆菌中有效的示例性启动子序列是T7启动子,但 是也可以使用在原核细胞中有功能的任何启动子或增强子。有用的启动子包括但不限于 lac启动子(大肠杆菌)、trp启动子(大肠杆菌)、araBAD启动子(大肠杆菌)、lac杂合启动子 (大肠杆菌)、trc杂合启动子(大肠杆菌)、PL(X)、SP6和T7。 启动子序列基于其在目标病原性细菌中实现可检测的表达水平的能力来选择。在 优选的实施方案中,报告基因优选与获自待检测的病原性细菌宿主的启动子偶联。无论生 理需求或底物浓度如何,组成型启动子以恒定速率表达报告体。或者,使用诱导型启动子以 控制报告基因表达的时机可能是有利的。对于诱导型启动子如lac和trp操纵子,表达通常 被阻遏并且可以在所需时间诱导。在不存在乳糖的情况下,lac启动子被lac阻遏蛋白阻遏。 诱导可以通过添加乳糖或IPTG来实现,从而阻止阻遏物与lac操纵子的结合。类似地,lip启 动子被与trp操纵子结合的色氨酸-阻遏物复合物负向调节。对于trp操纵子,可以通过去除 色氨酸或通过添加β-吲哚丙烯酸来诱导基因表达。 32 CN 111601897 A 说 明 书 30/43 页 (iii)细胞特异性复制起点 质粒中使用的复制起点可以是中等拷贝数,例如来自pBR322质粒(每细胞15-20个 拷贝)或R6K质粒(每细胞15-20个拷贝)的colE1 ori,或者它们可以是高拷贝数,例如,pUC oris(每细胞500-700个拷贝)、pGEM oris(每细胞300-400个拷贝)、pTZ oris(每细胞>1000 个拷贝)或pbluescript oris(每细胞300-500个拷贝)。复制起点可以在大肠杆菌或任何其 他原核物种如炭疽芽孢杆菌或鼠疫耶尔森氏菌(Yershinia pestis)中是功能性的。 质粒复制依赖于宿主酶及质粒编码的和质粒控制的顺式和反式决定子。例如,一 些质粒具有在几乎所有革兰氏阴性细菌中被识别且在复制起始和调控期间在每个宿主中 正确地起作用的决定子。其他质粒仅在一些细菌中具有这种能力(Kues等,Microbiol Rev 53:491-516,1989)。质粒通过三种通用机制复制,即θ型、链置换和滚环(由Del Solar等, Microbio and Mollec Biol Rev 62:434-464,1998综述)。 对于通过θ型机制的复制,复制起点可以定义为(i)可以支持质粒自主复制的最小 顺式作用区域,(ii)其中DNA链解链以启动复制过程的区域,或(iii)前导链合成开始处的 碱基。复制起点包含质粒和/或宿主编码蛋白质的相互作用所需的位点。经历θ型复制的质 粒还包括pPS10、RK2(含oriV)、RP4、R6K(含oriy)、ColE1和CoIE2。ColE1是通过θ型机制复制 的一类小型多拷贝质粒的原型。C61E1的复制起点跨越约1kb的区域(Del Solar等,1998)。 通过链置换机制的质粒复制的实例是IncQ家族的混杂质粒,其原型是RSF1010。该 家族的成员需要三种质粒编码的蛋白质来启动DNA复制。这些蛋白质促进复杂起源区域 (complex origin region)处的起始,并且复制通过链置换机制以任一方向进行。当RSF110 复制蛋白(RepA、RepB和RepC)由第二质粒以反式提供时,复制起点被定义为能够支持双向 复制的最小区域。最小ori区域包括三个相同的20-bp重复子加174bp区域,其含有富含GC的 延伸区段(28bp)和富含AT的区段(31bp)(Del Solar等,1998)。 通过滚环(RC)机制的复制是单向的,并且被认为是不对称过程,因为前导链的合 成和后随链的合成是解偶联的。关于RC复制的分子机制的研究主要用葡萄球菌质粒pT181、 pC221、pUBHO、pC194以及链球菌质粒pMV158及其Amob衍生物pLS1进行的。经历RC复制的其 他质粒或噬菌体包括但不限于pS194、fd、 pE194和pFX2(Del Solar等,1998)。 原核生物具有环状的染色体DNA分子,通常具有单一复制起点。例如,大肠杆菌和 其他细菌的染色体复制起点称为oniC。本方法设想使用本领域已知的从物种特异性质粒 DNA(例如,上文讨论的ColE1、R1、pT181等)、从噬菌体(例如 和M13)和从细菌染色体复 制起点(例如oriC)鉴定的复制起点。 (iv)抗生素抗性基因 转导颗粒的质粒DNA任选地具有抗生素抗性基因以促进质粒的分子生物学克隆并 允许选择由质粒转化的细菌。抗生素抗性基因是本领域公知的,且包括但不限于氨苄青霉 素抗性(Ampr)、氯霉素抗性(Cmr)、四环素抗性(Tetr)、卡那霉素抗性(Kanr)、潮霉素抗性 (hyg或hph基因)和吉欧霉素(zeomycin)抗性(Zeor)。优选地,抗生素抗性基因保护细菌免 受除正在测试其抗性或易感性的药物以外(或与其不同)的药物的抗微生物或细胞毒性作 用。在另一实施方案中,抗生素抗性基因保护细菌免受与正在测试其抗性或易感性的药物 相同的药物的抗微生物或细胞毒性作用。 F.制备转导颗粒的方法 33 CN 111601897 A 说 明 书 31/43 页 该方法中使用的转导颗粒或重组噬菌体通过修饰天然存在的噬菌体以携带能够 将目标细菌转化为易于识别的表型的基因(下文称为报告基因)而获得。转导颗粒必须能够 以使得细菌宿主可以以可检测的方式表达基因功能的方式将报告基因特异性地引入目标 细菌宿主中。存在大量噬菌体,并且可以基于所需的宿主范围和噬菌体携带和转导目标基 因的能力来选择进行修饰。特别地,噬菌体必须足够大以容纳报告基因、相关启动子区、噬 菌体包装位点和任何其他可包括的DNA区域。修饰的噬菌体通常保留未修饰的噬菌体的正 常宿主范围特异性,尽管特异性的一些改变是可接受的,只要它不影响鉴定所选目标细菌 的能力即可。 待修饰的噬菌体可以是温和的或毒性的,优选是温和的。噬菌体的修饰产生仍然 能够将报告基因引入靶细菌宿主中的转导颗粒。报告基因是质粒或其他自我复制的游离基 因单元的部分,其在感染的宿主中维持和表达。 报告基因的转导可以通过报告基因的瞬时表达(即,不是由细胞稳定遗传的报告 基因的表达)来进行。在这种情况下,通过噬菌体转导的DNA在整个测试期间可能不完整存 在。然而,由噬菌体转导的报告基因的转录在DNA被降解之前是足够有效的,以确保在测试 期结束时细菌组装了功能性报告基因。因此,即使细菌降解噬菌体DNA,细菌可以通过该测 定检测。 可用于该方法的噬菌体可以从微生物保藏库获得,例如美国典型培养物保藏中 心,P.O.Box 1549,Manassas,VA.,20108,USA。毒性噬菌体可作为无细菌裂解物获得,而溶 原性噬菌体通常可作为感染的宿主细胞获得。可以通过常规重组DNA技术修饰从任何来源 获得的野生型噬菌体,以引入能够产生可检测目标表型的所需报告基因。在引入之前,将目 标报告基因在合适基因盒上与启动子区域组合。基因盒可以通过常规重组DNA技术在合适 的宿主(如大肠杆菌)中构建。应选择报告基因和启动子区域以使两者在目标宿主中发挥功 能,并且该盒可任选地包括第二报告基因,如抗生素抗性、重金属抗性等,以利于体外操作。 报告基因(或多个基因,如果不是单基因系统)能够在目标细菌宿主中表达可筛选 的表型。如下文所用,短语可筛选表型意指允许表达该表型的细胞与不表达该表型的其他 细胞区分开的特征或性状,即使在所有细胞在混合培养中正常生长和繁殖。也就是说,特征 或性状的检测可以在感染的目标细胞存在于不表达该表型的存活的,通常增殖的非目标细 菌的混合群体中的同时进行。优选地,可筛选的表型包含可视觉检测的性状,即可以在目标 细胞和非目标细胞的混合群体中直接或间接观察到的性状。表型通常不是可选择的,即在 特定条件下提供存活或优先生长的表型(阳性选择)或在特定条件下提供生长抑制或杀灭 的表型。该方法不要求样品中存在的目标细菌与可能存在于样品中的非目标细菌分离或相 对于其富集,因为该性状在目标细菌仅包含存在的活细菌的一部分时是可观察到的。 报告基因可以自身编码可筛选的表型,或者可以是编码该表型的多基因系统的部 分,其中其他基因存在于待检测的宿主中或单独引入待检测的宿主中。例如,转导颗粒可携 带lacZa基因,其需要宿主中的互补lacZp基因或lacZAM15缺失用于表达。 合适的可筛选表型包括生物发光、荧光、酶催化的生色(例如,使用碱性磷酸酶) 等。可以通过常规可视化技术观察这些表型中的每一种,所述常规可视化技术提供完成信 号产生反应所必需的化学试剂。优选的是使用免疫检测,和更特别地用于检测异源酶或蛋 白质或者用于检测与酶或蛋白质共表达的分子的侧流免疫分析,其中共表达的分子用作功 34 CN 111601897 A 说 明 书 32/43 页 能性表达系统的指示物。 对于噬菌体,有可能通过将DNA构建体中噬菌体包装位点与质粒或盒连接来包装 质粒或报告基因盒。包装位点可以从噬菌体基因组获得并克隆到携带报告基因、启动子区 域和任选的第二报告体的质粒中。然后可以将质粒转移到合适的细菌宿主中。细菌宿主然 后产生具有包装在噬菌体衣壳内的质粒和/或标记基因盒的转导颗粒,其能够将质粒DNA插 入其宿主范围的细菌中。通过用质粒和噬菌体两者同时感染合适的宿主将质粒转入所需的 噬菌体中。孵育宿主细胞并收集转导颗粒。一部分噬菌体将携带质粒。可以通过常规技术将 转导颗粒与噬菌体分离。 宿主特异性噬菌体包装位点基本上和噬菌体基因组中与其相邻的天然序列分离。 如本文所用,关于噬菌体包装位点的术语“基本上分离的”是指它们不在其天然环境中。也 就是说,包装位点不是天然存在的全长噬菌体基因组核酸序列。包装位点可以通过实验技 术从全长噬菌体基因组序列分离,例如使用限制性核酸内切酶和通过聚合酶链式反应的克 隆或扩增。包装位点也可以合成产生。 噬菌体包装位点是全部或部分缺乏通常与其天然相关的序列的核酸片段,或者如 其天然存在的,但具有与其结合的异源序列的序列。它是从噬菌体基因组分离的片段。 如本文所用,在噬菌体包装位点的情况中的短语“功能等同物”是指定性地与野生 型噬菌体包装位点发挥相同功能的包装位点。因此,如果分离的噬菌体包装位点指导DNA的 包装,则如果DNA片段也以相同的方式指导DNA的包装,该DNA片段是功能等同物。根据该方 法,片段是功能等价物不需要量的等同。因此,具有核苷酸取代、缺失和/或添加的噬菌体包 装位点可以是分离的噬菌体包装位点的功能等同物。 G.使用噬菌体的方法 前述实施方案可以使用由完全完整的噬菌体或其包含最小结构元件的变体构成 的转导噬菌体颗粒来实施以允许将颗粒转导到宿主细胞中。在一些情况下,有可能感染生 物样品和直接观察改变和表型,但在其他情况下,可能优选首先制备样品中存在细菌的大 量培养物。用于获得样品和(如果需要)制备大量培养物的方法根据生物样品的性质而变 化,并且用于制备各种样品类型的合适技术在标准微生物学和细菌学教科书中详细描述, 例如Bergey 's Manual of Determinative Bacteriology(8th ed .),Buchanan and Gibbons(编辑)Williams&Wilkens Co .,Baltimore(1974);Manual of Methods for General Bacteriology,Gerhardt等(编辑),Am .Soc .Microbiology ,Wash .(1981);和 Ma n ua l of C l i n i ca l M i c r o bi o l o g y (8 t h ed .) ,Pa t r i c k ,R等 (ed s .) , Am.Soc.Microbiology,Washington(2003)。 噬菌体本身可以以各种形式添加到样品中。它可以以干燥状态添加。噬菌体可以 混合或悬浮在液体试剂混合物中。它可以悬浮在加入样品的小瓶中。它也可以采取任何其 他合适的形式。添加到样品中的噬菌体有时在本文中称为“亲本噬菌体”。一旦与噬菌体接 触,该样品被称为噬菌体暴露的样品。 噬菌体暴露的样品可以孵育预定的时间。如果存在于暴露的样品中,孵育可持续 足够的时间以允许在感染的目标细菌中产生噬菌体标志物。噬菌体暴露的样品处于有利于 目标细菌的噬菌体感染的条件中。这可以通过多种方式实现。例如,可以将亲本噬菌体混入 试剂中,其当添加到样品中时产生有利于感染的测试样品。可以以许多不同方式制备样品 35 CN 111601897 A 说 明 书 33/43 页 以建立有利于噬菌体感染的条件。 假定样品中存在目标细菌,测试样品将含有噬菌体标志物。亲本噬菌体通过将自 身附着于目标细菌的细胞壁并注入病毒核酸以产生感染的细菌的方式来感染目标细菌。重 组噬菌体标志物基因然后在感染的细菌中大量表达。如果细菌具有代谢活性,例如生长或 分裂,那么每个后代将含有标志物基因的额外拷贝(或被噬菌体感染),因此产生更大的信 号。相反,如果细菌静止或死亡,则产生较小的信号。 可以通过实施多个处理步骤来分析标志物。在一个实施方案中,该方法包括裂解 细菌。在一个实施方案中,微生物溶菌酶加入到噬菌体暴露的样品中。在一个实施方案中, 裂解包括向噬菌体暴露的样品加入氯仿、用酸处理噬菌体暴露的样品或以其他方式物理处 理噬菌体暴露的样品。 与其他方法相反,不需要产生后代噬菌体、宿主破裂、将子代噬菌体释放到测试样 品中以及随后的细菌感染轮次。此外,虽然许多现有技术方法依赖于检测完整的后代噬菌 体,但本公开的实施方案涉及检测过表达的标志物蛋白,其不是由细菌或噬菌体或细菌感 染的宿主例如人天然表达的。在其他实施方案中,标志物基因的产物可以赋予某些表型,例 如抗生素抗性或增强的生长性质,其可以在功能上筛选。 在一个实施方案中,噬菌体标志物是样品中目标细菌存在的间接指示物。在噬菌 体标志物是亲本噬菌体的组分的情况中,可以控制暴露样品中亲本噬菌体的初始浓度,使 得产生的背景信号在测定中是不可检测的。因此,如果样品中不存在目标细菌,则不发生感 染,不表达重组噬菌体标志物基因,并且不合成新的噬菌体标志物。在一个实施方案中,使 用已知缺乏目标细菌类型的对照样品进行阴性对照以确认所用的噬菌体在测定中不产生 背景信号。本公开的其他方面可以提供阴性对照用于鉴别可以与包含目标细菌的暴露样品 产生的任何信号区分开的背景信号。 在某些实施方案中,一旦制备了生物样品(有或没有大量培养物的生长),它通常 在促进颗粒与细菌的结合和遗传物质的注入的条件下不搅动地暴露于转导颗粒足够的时 间以允许感染(例如,15分钟至120分钟),通常在支持细菌快速生长的温度(例如,35℃至40 ℃)。感染后,孵育细胞以允许报告基因的表达,并且如下所述检测报告基因表达。 该方法适用于均质分离物以及异质细菌样品,包括例如多个细菌物种。术语“多 个”意指两个或更多个单元,例如细菌物种,尽管单个单元不需要在结构上和/或功能上不 同。在某些实施方案中,可以筛选样品以提供均质细菌群体,例如,使用适应于特定群体的 特定营养培养基。 与旨在产生转导细菌细胞的均质集落的常规噬菌体转导技术相反,该方法不需要 以任何方式分离转导的细菌。相反,可以在其中存在活的,通常是增殖的非目标细菌的生物 样品的未选择部分中检测由报告基因或其产物赋予的可筛选表型,例如可视觉观察的性 状。筛选可以在没有选择的情况下进行,因为不需要分离转导的细菌。 在一些实施方案中,该方法包括分析样品中标志物核酸或其产物的存在或不存 在。用于检测标志物核酸或其产物的合适方法是本领域已知的,并且可以和将根据样品的 性质而变化。 在一些实施方案中,用于确定细菌对抗生素的易感性或抗性的方法通过进行上述 抗生素处理、噬菌体转化和检测步骤来提供。抗生素处理和噬菌体转化的这些步骤可以以 36 CN 111601897 A 说 明 书 34/43 页 任何顺序进行或同时进行。在一个实施方案中,顺序进行抗生素处理和噬菌体转化的步骤。 本文所用的术语“顺序地”是指步骤按序进行,例如以数分钟、数小时、数天或数周的一个或 多个间隔进行。如果合适,步骤可以以规则的重复循环进行。在另一实施方案中,抗生素处 理和噬菌体转化步骤一起进行,然后进行测定步骤。 H.检测方法 检测报告基因或其产物的方法可以是间接的或直接的。间接检测可包括将报告基 因或其产物与样品中的其他组分分离,或将报告基因或其产物在样品中浓缩,然后在纯化 或浓缩的样品中检测报告基因或其产物。报告基因或其产物可以以释放的状态(例如,在含 噬菌体的培养基中)或以结合形式(例如,以在细胞溶质中或整合到基因组中的方式包含在 细菌内)检测。在一些情况下,报告体是在细菌表面上表达的分子,其允许在不需要裂解的 情况下进行其检测。在其他实施方案中,报告体可以是具有酶活性的蛋白质,例如CAT活性 或AP活性,如之前所述的。在这种情况下,使用酶促技术确定报告体活性。在又一个实施方 案中,报告体可以是对已知抗体或已知适体或蛋白质的结合配偶体显示出抗原性的蛋白 质。 在优选的实施方案中,报告体基因或其产物通过检测基因的蛋白质产物或蛋白质 片段(例如,含抗体与其特异性结合的抗原决定簇的肽)的存在来直接检测。在这一方面,表 位结合剂如抗体、适体或识别报告蛋白或其片段的其他分子配体可用于检测报告蛋白或其 片段。在一个示例性实施方案中,抗体或其抗原结合片段用于检测报告蛋白或其片段的存 在。在其他实施方案中,抗体可用于检测由报告蛋白的生物合成活性产生的产物,例如,其 中报告体是具有针对另一蛋白质的特异性活性的蛋白酶,检测第二蛋白质的消化产物。 在一个实施方案中,使用质谱法检测报告蛋白或其片段。特别地,可以使用将色谱 步骤与质谱步骤连接的技术。一般而言,报告蛋白或其片段的存在可以使用液相色谱然后 质谱法来确定。 在一些实施方案中,液相色谱法是高效液相色谱法(HPLC)。HPLC的非限制性实例 可包括分配色谱、正相色谱、置换色谱、反相色谱、尺寸排阻色谱、离子交换色谱、生物亲和 色谱、水性正相色谱或超快速液相色谱。在一个实施方案中,液相色谱可以是超快速液相色 谱。 在一些实施方案中,质谱可以是恒定中性丢失质谱。在其他实施方案中,质谱可以 是串联质谱法(MS/MS)。在不同的实施方案中,质谱可以是基质辅助激光解吸/电离 (MALDI)。在具体实施方案中,质谱可以是电喷雾电离质谱(ESI-MS)。 在示例性实施方案中,该方法包括液相色谱,接着串联质谱。在特别示例性的实施 方案中,该方法包括如实施例中所述的液相色谱接着串联质谱。在另一示例性实施方案中, 该方法包括液相色谱接着恒定中性丢失质谱。在特别示例性的实施方案中,该方法包括液 相色谱,然后如实施例中所述的恒定中性损失质谱。在又一个示例性实施方案中,该方法包 括液相色谱接着电喷雾电离质谱(ESI-MS)。 在每个上述实施方案中,液相色谱接着质谱可用于确定样品中报告蛋白或其片段 的存在,或者液相色谱接着质谱可用于确定样品中报告蛋白或其片段的存在和量。在优选 的实施方案中,液相色谱接着质谱可用于确定样品中报告蛋白或其片段的存在。 一般地,评估报告蛋白或其片段的存在或量的基于表位结合剂的方法包括在有效 37 CN 111601897 A 说 明 书 35/43 页 地允许形成表位结合剂和报告蛋白或其片段之间的复合物的条件下,使包含报告蛋白或其 片段的样品与对报告蛋白或其片段特异性的表位结合剂接触。基于表位结合剂的方法可以 在溶液中进行,或者表位结合剂或样品可以固定在固体表面上。合适的表面的非限制性实 例包括微量滴定板、试管、珠子、树脂和其他聚合物。 如本领域技术人员所理解的,表位结合剂可以广泛的方式附接于基质。表位结合 剂可以首先合成,随后附接到基质上,或者可以直接在基质上合成。基质和表位结合剂可以 用化学官能团衍生化以随后连接两者。例如,基质可以用化学官能团衍生化,包括但不限于 氨基、羧基、氧代基团或硫醇基。使用这些官能团,表位结合剂可以使用官能团直接连接或 使用接头间接连接。 表位结合剂也可以非共价地附接于基质上。例如,可以制备生物素化的表位结合 剂,其可以与链霉亲和素共价包被的表面结合,从而导致附接。或者,可以使用诸如光聚合 和光刻的技术在表面上合成表位结合剂。将表位结合剂连接到固体表面的另外的方法和在 基质上合成生物分子的方法是本领域公知的,即来自Affymetrix的VLSIPS技术(例如,参见 美国专利号6,566,495,和Rockett等,Xenobiotica 30(2):155-177,2000)。 为了允许在表位结合剂和报告蛋白或其片段之间形成复合物,将单个组分在有效 条件下孵育足够长的时间以使表位结合剂与存在的任何抗原结合。之后,复合物可以洗涤 以除去/减少非特异性结合,并且复合物可以通过本领域熟知的任何方法检测。检测表位结 合剂和报告蛋白或其片段之间的复合物的方法通常基于标记或标志物的检测。如本文所 用,术语“标记”是指附接于表位结合剂或其他基质材料的任何物质,其中该物质可通过检 测方法检测。合适的标记的非限制性实例包括发光分子、化学发光分子、荧光染料、荧光猝 灭剂、有色分子、放射性同位素、闪烁体、生物素、抗生物素蛋白、链霉亲和素、蛋白A、蛋白G、 抗体或其片段、聚组氨酸、Ni2 、Flag标签、myc标签、重金属和酶(包括碱性磷酸酶、过氧化 物酶和荧光素酶)。这些方法在本领域中是众所周知的。 在一些实施方案中,通过免疫分析检测复合物。免疫分析可以以多种不同的形式 进行。一般而言,免疫分析可分为两类:竞争性免疫分析和非竞争性免疫分析。在竞争性免 疫分析中,样品中未标记的分析物与标记的分析物竞争结合表位结合剂如抗体。洗去未结 合的分析物并测量结合的分析物。在竞争性免疫分析的可选实施方案中,样品中未标记的 分析物从固定的分析物置换标记的表位结合剂如抗体。测量置换的抗体的量作为样品中分 析物的量的指示。在非竞争性免疫分析中,表位结合剂(如抗体)而不是分析物被标记。非竞 争性免疫分析可以使用一种抗体(例如,捕获抗体被标记)或多于一种抗体(例如至少一种 未标记的捕获抗体和至少一种标记的“加帽”或检测抗体)。合适的标记以上描述。 在一些实施方案中,基于表位结合剂的方法是ELISA。在其他实施方案中,基于表 位结合剂的方法是放射免疫分析。在再其他的实施方案中,基于表位结合剂的方法是免疫 印迹或蛋白质印迹。在不同的实施方案中,基于表位结合剂的方法是免疫组织化学(IHC)。 在可选实施方案中,基于表位结合剂的方法是阵列。在不同的实施方案中,基于表位结合剂 的方法是侧流分析。侧流分析可以是旨在检测样品中靶分析物的存在(或不存在)的装置。 用于提高特异性鉴定细菌宿主的能力的第二种方法涉及使用免疫吸附。固定的抗 体,包括抗血清或单克隆抗体,用于基于其细胞表面表位的特性特异性地捕获细菌细胞。然 后可以使用转导颗粒进一步检测细菌,如上所述。用于在固相表面上对特定细菌物种和菌 38 CN 111601897 A 说 明 书 36/43 页 株进行免疫吸附的合适材料和方法描述于Enterobacterial Surface Antigens:Methods for Molecular Characterization,Korhonen等(编辑),Elsevier Science Publishers, Amsterdam(1986)中。 [0200] 在另一实施方案中,可以使用细胞计数技术检测报告蛋白或其片段。尽管在别处 已经报道了用于进行培养细菌的细菌计数测量的方法(Martinez等,Cytometry(1982)3 (2):129-33;Suller等,J Antimicrob Chemother(1997)40(1):77-83;和Roth等,Appl Environ Microbiol(1997)63(6):2421-31),但是它们不涉及噬菌体报告蛋白的检测。细胞 计数检测方法可适应于革兰氏阳性和革兰氏阴性细菌两者,例如大肠杆菌(Martinez,同 上)、蜡状芽孢杆菌(Roth,同上)、金黄色葡萄球菌(Suller等,J Antimicrob Chemother (1997)40(1):77-83)、表皮葡萄球菌(Cohen等,J Clin Microbiol(1989)27(6):1250-6)、 化脓性链球菌(Cohen,同上)、肺炎克雷伯菌(Cohen,同上)、铜绿假单胞菌(Cohen,同上)、施 氏假单胞菌(Cohen,同上)、奇异变形杆菌(Cohen,同上)和肠杆菌属(Cohen,同上)。 [0201] 在前述实施方案中,该方法利用宿主特异性重组或工程化噬菌体。例如,能够感染 鼠疫耶尔森氏菌的遗传修饰的 可用于特异性检测鼠疫耶尔森氏菌。为了检测多种靶 细菌类型,可以将对每种目标细菌类型特异性的一种噬菌体添加到单个测试样品中,或单 独地添加到其分部中。 [0202] 图1示出了根据本文描述的方法的一个实施方案的示例性工作流程。获得包含细 菌的样品10。如上所述,样品可以来自受试者、来自食品、来自环境等。如果需要或期望,可 以对样品进行加工或处理。等份的样品在抗生素存在的情况下孵育或培养12,并且任选地, 等份的样品在不存在抗生素的情况下孵育或培养。同时或顺序地,样品等分试样与对样品 中的细菌特异性的重组或工程化噬菌体一起孵育或培养12。如上所述,工程化噬菌体包含 异源标志物。然后分析14通过与抗生素和工程化噬菌体一起孵育产生的培养物以确定标志 物的存在或不存在(定量或定性),并报告结果或数据16。 [0203] 图2示出了根据本文描述的方法的另一实施方案的示例性工作流程。获得包含细 菌的样品20。如果需要或期望,样品可以加工或处理。制备包含具有和不具有抗生素的流体 培养基的容器22。等份的样品24放入每个容器中,并混合26。然后,将每个容器在所需温度 下孵育所需时间,并且在该实施方案中,将每个容器在35℃下孵育2小时(28)。容器可以在 孵育后再次混合,然后将工程化的噬菌体引入容器中,混合(34),然后孵育(36)以产生次级 培养物。将等份的次级培养物沉积在侧流免疫分析装置上(38),然后分析(40)异源标志物 的存在或不存在(定量或定性)。 [0204] 图1和2的工作流程是用于进行本文所述方法和试验以筛选候选抗生素对抗细菌 样品、细菌菌株或细菌菌株混合物的效力的示例。通常,该方法包括使细菌样品与测试抗生 素接触以获得原代培养物及与缺乏测试抗生素的媒介接触以获得对照原代培养物;使包含 编码异源报告基因表达的核酸序列的特定噬菌体与原代培养物和与对照原代培养物接触, 以获得包含用测试抗生素处理的细菌的第一次级培养物和包含未用测试抗生素处理的细 菌的第二次级培养物;和检测第一和第二次级培养物中报告基因或其产物的水平或活性, 其中第一次培养物中报告基因或其产物的水平或活性与第二次级培养物相比的降低(或不 存在)表明试验化合物是抗生素剂。该方法还用于筛选针对多种细菌菌株的单一测试抗生 素。该方法还用于抗生素或候选抗生素的最小抑制浓度(MIC)和/或筛选以确定临床抗生素 39 CN 111601897 A 说 明 书 37/43 页 化合物的功效。 [0205] 如本文所用,术语“最小抑制浓度”是指抑制微生物可见生长的抗生素的最低浓 度。该术语还包括使用本文所述的方法和试验影响细菌细胞死亡或抑制细胞壁修复的抗生 素的最低浓度。在一个实施方案中,本文所述的方法和试验允许测定抗生素或候选抗生素 针对细菌菌株的最小抑制浓度。在一个实施方案中,抗生素的最小抑制浓度可以通过测量 暴露于抗生素的样品中的细菌细胞的响应与未暴露于抗生素的样品中或暴露于不同浓度 的相同抗生素的样品中的相同细菌细胞相比的调节(例如,报告染料的摄取或排出、形态变 化、代谢变化等)来确定。 [0206] 最小抑制浓度是指示待施用于受试者以诱导细菌细胞死亡和/或减轻细菌介导的 疾病的至少一种症状的抗生素最小有效剂量的临床相关值。临床上,最小抑制浓度不仅用 于确定受试者将接受的抗生素量,而且还用于确定要使用的优选抗生素。还可以确定候选 抗生素的最小抑制浓度以允许例如功效测定和临床试验的给药信息。 [0207] 本方法可用于患者诊断,因为它们允许确定细菌对抗生素和其他杀菌剂的敏感 性。通过在暴露于转导颗粒之前进行细菌与待筛选抗生素或杀菌剂的短暂孵育,细胞的代 谢活性停止并防止表型的改变。在最初使用如上所述的转导颗粒确认细菌的存在后,这种 测试将是有用的。然后可以将确定对细菌感染致死的抗生素和杀菌剂用于治疗受试者。有 用的抗生素和杀菌剂的这种快速和早期检测在治疗严重的细菌感染中是非常有价值的。 [0208] 在一个实施方案中,诊断方法可包括使患有或怀疑具有细菌性疾病的风险的受试 者的样品与本文所述的一种或多种重组噬菌体接触;检测并任选地定量噬菌体表达的标志 物的存在或不存在;将标志物的存在与细菌性疾病的致病因子(例如,金黄色葡萄球菌)相 关联,从而诊断受试者中的细菌性疾病。“受试者”是指脊索动物门的任何成员,包括但不限 于人和其他灵长类动物,包括非人灵长类动物,如黑猩猩和其他类人猿和猴子物种;农场动 物,如牛、绵羊、猪、山羊和马;家养哺乳动物,如狗和猫;实验动物,包括啮齿动物如小鼠、大 鼠和豚鼠;鸟类,包括家禽、野生和猎禽,如鸡、火鸡和其他鹑鸡类、鸭、鹅等。该术语涵盖成 体和新生个体。 [0209] 在某些实施方案中,在进行细菌性疾病的阳性诊断后,受试者可以任选地根据标 准临床程序进行治疗、管理和随访。例如,受试者可以用有效量的药剂例如抗生素治疗。出 于本方法的目的,药剂的“有效量”是产生针对受试者中的致病因子(例如,金黄色葡萄球 菌)的响应的量。在这一方面,患有咽炎的受试者可以用青霉素G苄星青霉素和/或阿莫西林 治疗。如果发现受试者对治疗没有反应,可以使用上述方法分析致病因子的抗生素抗性。如 果对抗性菌株作出阳性鉴定,则受试者可以用第二抗生素剂或更高剂量的抗生素剂或者两 种或更多种抗生素剂的组合治疗。 [0210] 类似地,本方法可用于检测抗生素的存在,例如动物产品中的抗生素残留。在该方 法中,待分析材料的提取物添加到对所述抗生素敏感的细菌菌株的培养物中,将混合物孵 育预定时间段,如果存在给定量的抗生素该时间段足以杀死该菌株。此时,添加对菌株特异 性的转导颗粒,并进行如本文所述的分析。如果存在给定量的抗生素,则细菌菌株的细胞将 死亡并且读数将是阴性的(即,荧光素酶试验中缺乏发光)。如果给定量的抗生素不存在或 低于MIC,则细菌将存活并且读数将是阳性的(即发光)。 [0211] 在具体实施方案中,提供了一种用于测定先前已经赋予噬菌体特异性的对转导颗 40 CN 111601897 A 说 明 书 38/43 页 粒敏感性的细菌的方法。即,在第一步中,修饰目标细菌(例如通过转化),使得它们含有或 表达目标噬菌体的细胞特异性受体。在第二步中,修饰的(或标记的)细菌引入或混合到其 中它们将要被持续关注的样品环境中。样品环境可以是其中存在细菌的任何环境,包括室 外(例如土壤、空气或水);在存活宿主(例如植物、动物、昆虫)上;在设备(例如,制造、加工 或包装设备)上;和临床样品中。然后使用对引入的受体特异性的噬菌体进行如本文所述的 噬菌体试验,并且可以监测或定量标记的细菌的存在。 [0212] 参照以下非限制性实施例进一步描述前述实施方案。 实施例 [0213] 本文描述的结构、材料、组合物和方法旨在作为本发明的代表性实例,并且应理 解,本发明的范围不受实施例的范围的限制。本领域技术人员将认识到,可以对所公开的结 构、材料、组合物和方法进行变化来实施本发明,并且这些变化被认为是在本发明的范围 内。 实施例1 用于表达异源标志物的重组噬菌体的构建 [0214] 在本研究中,制备不是由靶细胞或由噬菌体载体或由细菌感染的宿主天然产生的 分子(标志物),然后通过免疫分析特异性地检测异源分子(标志物)。标志物分子可以是肽 或蛋白质,任选地用例如聚组氨酸(His)标签等标记。 [0215] 重组噬菌体的构建:重组噬菌体经由重组质粒介导的同源重组通过将编码异源标 志物的DNA序列插入编码衣壳蛋白(cps)的核酸序列下游的噬菌体基因组的强表达区域中 来构建。位于cps上游的强启动子在感染后噬菌体基因组表达过程中被选择性激活,从而产 生相应mRNA转录物的许多拷贝。使用编码报告体的核酸的融合产物完成重组噬菌体的构 建,其具有合适的翻译信号(核糖体结合位点、中间区域、起始密码子)。代表性方法在 Loessner等,Appl.Environ.Microbiol.,62(4):1133-40,1996和美国专利No.5,824,468中 描述。 [0216] 质粒载体电转化到电感受态大肠杆菌K1菌株(ATCC菌株23503)中的电转化:通过 在Luria-Bertani(LB)肉汤中于37℃下生长至0.8的光密度(OD),然后在15%甘油中洗涤几 次使菌株成为电感受态。电转化用可获自Bio-Rad Labs(Hercules,CA)的GENE PULSER完 成。 [0217] 转化的大肠杆菌K1菌株用原生型噬菌体的感染:宿主细菌感染后,至少少数原生 噬菌体与质粒中侧邻luxAB的衣壳序列的部分进行同源重组,因此转移luxAB以形成重组噬 菌体。转化的细菌在37℃下在LB-氨苄青霉素培养基中生长至OD为0.4。噬菌体K1-5以每10 个细菌约1个噬菌体的感染复数(MOI)加入,并监测OD直至发生裂解。通过0.45微米硝酸纤 维素膜(可获自Millipore Corp.,Bedford,MA)过滤来收集裂解产物。 [0218] 将裂解产物铺板并使用连续稀释在生长于含有50μg/mL氨苄青霉素的LB固体琼脂 上生长的野生型大肠杆菌K1(ATCC 23503)上形成斑块(plaqued)和通过测定噬斑以确定报 告体活性来筛选重组K1-5噬菌体。可以通过对噬菌体基因组进行测序来确认已经产生含有 适当整合的报告基因序列的重组噬菌体。借助商业测序仪 (C o m m o n w e a l t h Biotechnologies,Richmond,VA)完成测序。 41 CN 111601897 A 说 明 书 39/43 页 [0219] 最后,通过免疫分析检测表达的标志物。 实施例2 使用DNA转位产生重组噬菌体 [0220] 使用商业上可获得的EZ::TNTM转座酶系统(Epicenter Technologies,Madison, WI)构建含有异源报告核酸的噬菌体,如Goryshin等,J.Biol.Chem.,273(13):7367-7374, 1998中所述。然后,将末端ME结合的EZ::TNTM转座体电转化到电感受态大肠杆菌K1菌株 (ATCC菌株23503)中。通过在LB培养基中于37℃下生长至0.8的光密度(OD),然后在15%甘 油中洗涤数次使菌株成为电感受态。使用获自Bio-Rad Laboratories(Hercules,CA)的 BIORAD GENE PULSER完成电转化。 [0221] 转化的大肠杆菌K1菌株用原生型K1-5噬菌体感染。转化的细菌在37℃下在LB-氨 苄青霉素培养基中生长至OD为0.4。噬菌体K1-5以每10个细菌约1个噬菌体的感染复数加 入,并监测OD直至发生裂解。在转座体-电转化的宿主细菌的感染后,至少少数原生K1-5噬 菌体通过在不影响噬菌体的噬斑形成能力的无害位置处的随机转座接受异源报告基因。通 过0.45微米硝酸纤维素膜(获自Millipore Corp.,Bedford,MA)过滤来收集裂解产物。 [0222] 使用标准方法筛选裂解产物的报告基因产物的活性,例如使用特异性结合报告蛋 白的抗体或片段的对于报告蛋白的免疫分析。可以用含有可检测部分的第二抗体检测第一 抗体。 实施例3 使用重组噬菌体筛选从受试者获得的样品 [0223] 鉴定出表现出细菌感染的症状(例如,发烧、头痛、腹痛和恶心)的受试者(例如,人 类患者),并且从受试者收集以下样品:0.01mL脑脊液(CSF)样品,1.0mL痰液样本和1.0mL血 液样本。各样品用4.0mL LB肉汤稀释,从而促进相应样品中存在的所有细菌的生长,并在37 ℃下孵育4小时。孵育后,将每个样品以100μL等分试样分配到96孔板的30个孔中。将等份的 血液样品加入孔1-30中,将等份的CSF样品加入孔31-60中,并将等份的痰液样品加入孔61- 90中。孔91-93用作阳性对照,且孔94-96用作阴性对照。 [0224] 获得以下五种重组噬菌体:K1-5::luxAB噬菌体,其感染大肠杆菌K1细菌;EBN6:: luxAB噬菌体,其感染肠球菌;Twort::luxAB噬菌体,其感染葡萄球菌;Sp6::luxAB噬菌体, 其感染沙门氏菌;和RZh::luxAB噬菌体,其感染链球菌。使用实施例1中描述的方案,噬菌体 从另一来源获得或原位工程化。 [0225] 将相当于约3×108噬菌体/mL的重组噬菌体悬浮液加入到对应于从患者收集的三 个样品中每一个的30个孔的组的6个单独孔中。由于该系统涉及在不同特异性的噬菌体上 应用相同的异源标志物,因此开发了通用检测系统。通过微小调整,该系统适应于多重检 测,例如,其中多个样本一起或同时处理。后一系统特别可用于鉴定参与特定疾病发病机理 的细菌病原体群。例如,与尿路感染(UTI)相关的病原体,例如大肠杆菌、克雷伯氏菌、肠杆 菌、假单胞菌、葡萄球菌、变形杆菌,都可以使用多重阵列形式进行鉴定和表征。 实施例4 确定细菌样品的抗微生物剂易感性或抗性 [0226] 生物样品用4.0mL LB肉汤稀释,并在37℃下孵育4小时以促进样品中所含细菌的 生长。将原代培养物均匀等分到含有补充有以下测试抗生素物质之一的LB培养基的 42 CN 111601897 A 说 明 书 40/43 页 Eppendorf管中:氨苄青霉素(青霉素类)、亚胺培南(β-内酰胺类抗生素)、头孢西丁(头霉素 类)、环丙沙星(氟喹诺酮类)、卡那霉素(氨基糖苷类)和四环素(四环素类)。在用抗生素处 理后,将管离心,且洗涤细菌沉淀并重悬于包含含有报告基因的噬菌体颗粒的LB肉汤中。噬 菌体-细菌混合物在标准条件下培养以允许细菌的转化和次级培养物的产生。将次级培养 物离心,并用裂解缓冲液裂解细菌沉淀。使用标准技术测定报告基因产物(异源蛋白质)的 量或活性,例如,ELISA以评估报告异源蛋白的量或酶分析以评估报告基因产物(异源蛋白) 的活性。该实验可以以单重或多重形式进行。 [0227] 单重测定法和多重测定法的基本概述在下表4.1中给出。预期结果在表4.2中给 出。 表4.1:基本实验设置(多重形式为粗体) 表4.2:免疫分析的预期结果(多重形式为粗体) 43 CN 111601897 A 说 明 书 41/43 页 实施例5 抗细菌剂易感性测试方法 [0228] 保留实施例4的基本实验设置,除了用增加的对数浓度的相同抗生素处理多批测 试样品,例如,终浓度0.00mg/L(对照)、0.01mg/L、0.03mg/L、0.10mg/L、0.30mg/L、1.0mg/L、 3.0mg/L、10mg/L、30mg/L、100mg/L氨苄青霉素。对于每种测试的单个测试化合物,确定针对 每一单个细菌物种的氨苄青霉素的最小抑制浓度(MIC)曲线。可以比较彼此相关的物种或 菌株,例如,关于抗生素的50%致死剂量(LD50)和/或细菌的MIC值以及生长参数,例如抗生 素对倍增时间的影响,等。 实施例6 苏云金芽孢杆菌Cry1Ab毒素的检测方法 [0229] 使用市售抗体制备用于检测作为标志物蛋白的苏云金芽孢杆菌(Bt)的Cry 1Ab蛋 白毒素的侧流免疫分析,以检测纯化蛋白质形式的Cry1Ab和来自裂解的大肠杆菌细胞的质 粒编码蛋白质。评估该方法直接鉴定来自尿液的病原体并确定其抗微生物剂易感性(ID/ AST)的能力,并与为抗微生物剂易感性测试(AST)设计的临床和实验室标准协会(Clinical and Laboratory Standards Institute,CLSI)指南提供的标准方案进行比较。 [0230] 使用商业抗Bt Cry1Ab兔多克隆抗体和鼠单克隆抗体(MyBioSource)制备侧流免 疫分 析 。使 用侧流免 疫分 析 检 测 C r y 1 A b 与使 用市 售 E L I S A 试 剂盒 ( A M A R Immunodiagnostics,目录号AID007)的Cry1Ab检测进行比较。侧流免疫分析具有通用捕获 和检测试剂以允许有效筛选标志物特异性的抗体。捕获试剂由固定在支持膜上的抗荧光素 单克隆抗体组成。检测试剂是链霉亲和素包被的铕纳米颗粒(ThermoFisher)。标志物特异 性的抗体使用商业试剂(异硫氰酸荧光素和生物素-NHS酯)用荧光素和生物素标记,并以侧 流免疫分析的形式测试与纯化的Bt Cry1Ab蛋白的配对。关键的免疫分析参数-捕获和检测 试剂的浓度、分析缓冲液和阻断剂的组成、样品体积-用来自MyBioSource的纯化的Bt Cry1Ab蛋白(UniProt P0A370)优化。 [0231] 评估了侧流免疫分析检测携带Cry1Ab蛋白表达载体(pCiylAb-ET30a)的大肠杆菌 44 CN 111601897 A 说 明 书 42/43 页 细胞产生的Cry1Ab蛋白的能力。测试系统用纯化的Bt Cry1Ab蛋白(MyBioSource# MBS537737)校准,且检测限使用纯化蛋白的标准曲线计算为对应于[空白样品信号 3SD]的 信号的Cry1Ab蛋白浓度。 [0232] 评估了大肠杆菌细胞中的Cry1Ab蛋白表达、其从裂解细胞的细胞外释放和CFU/mL 检测限。合成编码Cry1Ab的基因(Epoch Life Sciences)并亚克隆到pET30a(Novagen)中。 pET30a是在T7 RNA聚合酶1的转录控制下的蛋白质表达载体。将pCrylAb-ET30转化到大肠 杆菌B中并用T7噬菌体感染。T7感染诱导Cry1Ab表达(通过提供RNA聚合酶1),并且在感染后 30-45分钟之后,裂解释放Cry1Ab的细胞。使用侧流免疫分析确定噬菌体诱导的Cry1Ab表达 的存在和从裂解细胞的释放。检测能力使用范围为102至106CFU/mL,恒定108PFU/mL的细胞 连续稀释评估。由于只有噬菌体感染的裂解细胞在上清液中产生Cry1Ab,这种使用噬菌体 诱导的质粒产生Cry1Ab表达的策略为后续实验提供了有用的基准。 [0233] 其它实施方案:前述实施例可以通过将本说明书中其他地方描述的一般或具体描 述的反应物和/或操作条件替换为前述实施例中使用的那些重复而获得类似的成功。 实施例7 尿道感染的检测方法 [0234] 在本实施例中描述了用于诊断尿路感染的诊断分析,其鉴定存在的细菌。该分析 包括i)用于识别目标细菌的重组噬菌体和使用表达异源蛋白质标志物的目标细菌,和ii) 用于检测异源蛋白质标志物的侧流免疫分析。该分析考虑用于临床样品,例如尿液,其与含 有抗生素和重组噬菌体的试剂一起孵育以形成混合物;在孵育后,将混合物应用于侧流免 疫分析以确定异源标志物的存在或不存在。抗生素敏感的细菌菌株被抗生素抑制和不表达 标志物,且侧流免疫测分析不产生信号(阴性结果),而抗生素抗性细菌菌株产生标志物且 导致侧流免疫分析的可检测信号(阳性结果)。 [0235] 作为表达组氨酸标记的生物素化荧光素酶作为异源蛋白标志物的重组噬菌体的 模型,制备表达具有生物素或生物素样部分(链霉亲和素结合蛋白(SBP)和 )的 H i s -标记的荧光素酶构建体的重组大肠杆菌细胞。这六种重组细胞系命名为 HM50NanoSBPHis、HM50NanoHisSBP、HM50HisTwinStrept、HM50TwinStreptHis、 HM50HisBiotin、HM50BiotinHis。用Y-PER Plus蛋白质提取试剂从每个细胞系制备裂解产 物。 [0236] 侧流免疫分析被构建,具有在硝酸纤维素基质上以上游至下游位置的以下区域: 具有分别针对His标签的鼠抗体和链霉亲和素包被的铕颗粒的捕获和检测试剂的试剂垫; 由山羊抗体的阴性对照线、具有针对鼠IgG的山羊抗体的测试线和生物素化的牛血清白蛋 白(BSA)的程序控制线组成的测试区;和吸收垫。 [0237] 将细胞裂解产物的稀释提取物置于侧流免疫分析上,并用仪器读取测试区中的线 处的信号。从命名为HMSOBiotinHis、HM50HisBiotin和HM50NanoHisSBP的细胞的裂解产物 观察到可检测信号。表达HM50HisBiotin的细胞提供了最高的可检测his-荧光素酶-生物素 信号量。 [0238] 从前面的描述,本领域技术人员可以容易地确定该方法的必要特征,并且在不脱 离其精神和范围的情况下,可以进行各种改变和修改以使其适应各种用途和条件。 [0239] 除非另外定义,否则本文使用的所有技术和科学术语具有与本发明所属领域的普 45 CN 111601897 A 说 明 书 43/43 页 通技术人员通常理解的含义相同的含义。虽然与本文描述的那些类似或等同的方法和材料 可用于本发明的实施或测试,但在前面的段落中描述了合适的方法和材料。另外,材料、方 法和实施例仅是说明性的而非限制性的。如果冲突的话,以本说明书(包括定义)为准。 [0240] 本文引用的所有美国专利和公开或未公开的美国专利申请通过引用结合到本文 中。本文引用的所有公开的外国专利和专利申请均由此通过引用并入。本文引用的所有公 开的参考文献、文档、手稿、科学文献均由此通过引用并入。与科学数据库(例如,NCBI、 GENBANK、EBI)有关的所有标识符和登录号由此引入作为参考。 46 CN 111601897 A 说 明 书 附 图 1/2 页 图1 47 CN 111601897 A 说 明 书 附 图 2/2 页 图2 48