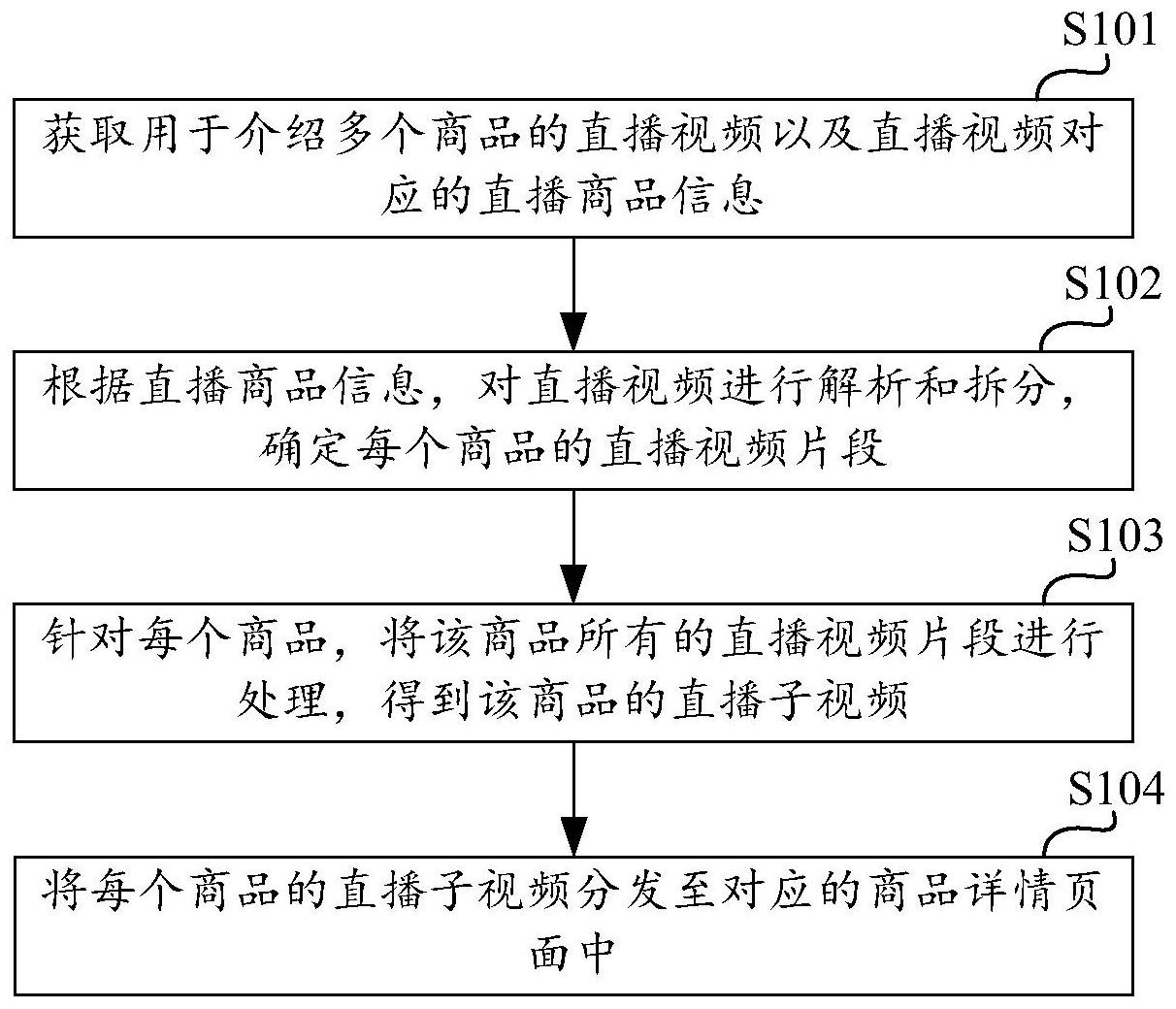
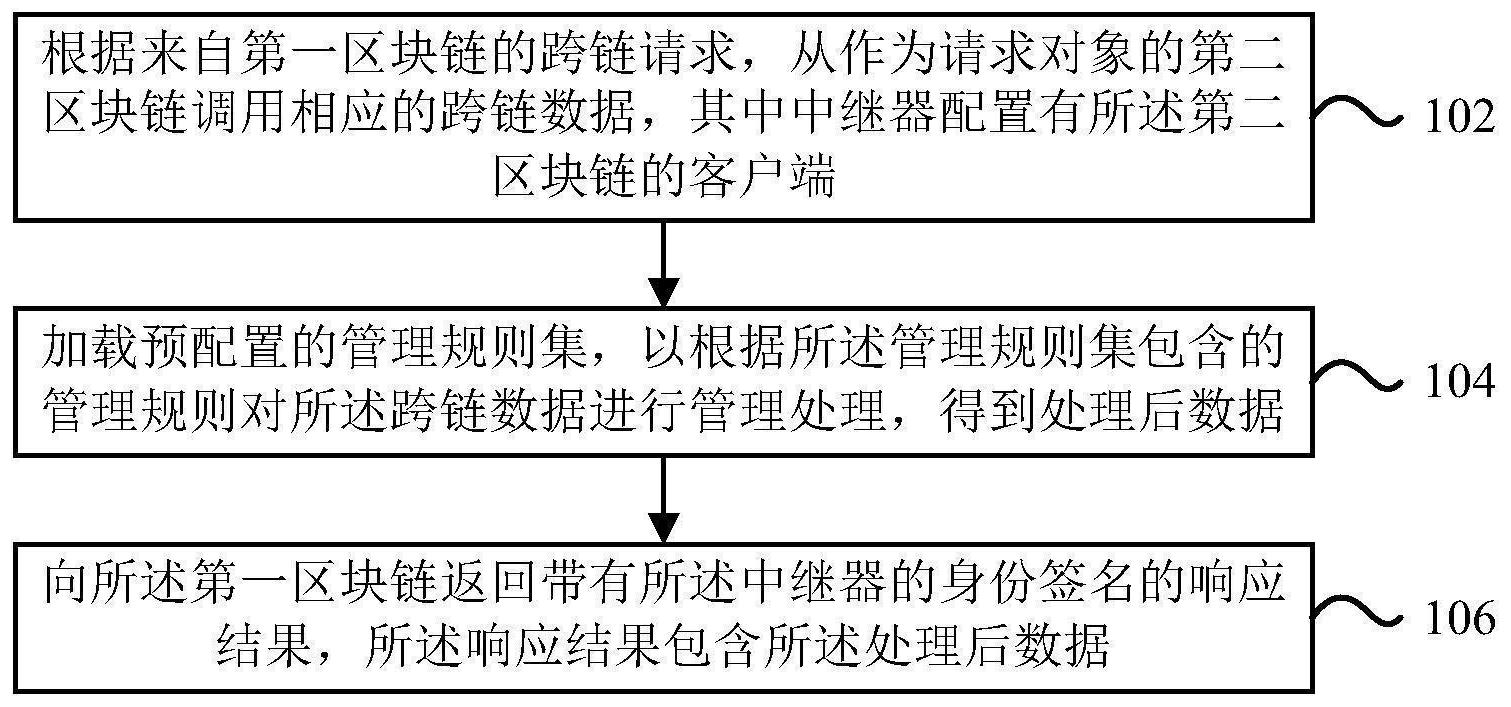
技术摘要:
本发明涉及腺相关病毒(AAV),其包括在对应于SEQ ID NO:1的第87和588位的位置之间的至少6‑8个氨基酸的插入物。还预期用作药物的本发明的AAV以及包含本发明的AAV的药物组合物。此外,本发明涉及本发明的AAV用于转导视网膜细胞核的体外用途。还涉及用于筛选插入序列的方 全部
背景技术:
重组腺相关病毒(AAV)载体已被证明是一种非常适合将基因高效且长期转移到视 网膜细胞中的递送系统(Boye et al.(2013)“A comprehensive review of retinal gene therapy”Molecular therapy ,21(2013)509-519and Trapani et al .(2014)“Vector platforms for gene therapy of inherited retinopathies”Prog Retin Eye Res)。AAV 是属于细小病毒科和依赖病毒属的非致病性病毒,仅在存在腺病毒、乳头状瘤病毒或疱疹 病毒的情况下才能复制(Nathwani et al.(2014)“Long-term safety and efficacy of factor IX gene therapy in hemophilia B”N Engl J Med ,371(2014)1994-2004),并具 有大约5kb单链DNA基因组(Trapani et al.(2014)“Vector platforms for gene therapy of inherited retinopathies”Prog Retin Eye Res.)。 AAV的安全性和转基因的长期表达已在大型动物模型、非人灵长类动物和多项人 类试验中被广泛测试(MacLaren et al .(2014)“Retinal gene therapy in patients with choroideremia:initial findings from a phase 1/2clinical trial”Lancet383, 1129-1137 ,Maguire et al .(2008)“Safety and efficacy of gene transfer for Leber 's congenital amaurosis”N Engl J Med ,358 2240-2248 ,Simonelliet al . (2010)”Gene therapy for Leber's congenital amaurosis is safe and effective through1.5years after vector administration”Molecular therapy:the journal of the American Society of Gene Therapy 18,643-650,Nathwani et al.(2014)“Long- term safety and efficacy of factor IX gene therapy in hemophilia B”N Engl J Med 371,1994-2004)。 AAV是用于基因补充疗法(也称为基因增强疗法)的优选基因转移载体。基因补充 疗法是用于治疗遗传疾病的有吸引力的方法,其目的在于恢复由致病突变引起的基因功能 不足或缺乏。目标是恢复受影响的细胞类型的正常生理功能和/或通过补充基因功能来抑 制或延迟变性过程。 基因疗法的目的是恢复由致病突变引起的基因功能不足或缺乏。这种方法对于隐 性遗传疾病的治疗特别有吸引力,在隐性遗传疾病中,异常转录物或蛋白质预计不会干扰 治疗。目标是通过在适当的位置补充足量的正常基因功能来阻止或防止变性过程,并恢复 受影响细胞类型或组织的正常生理功能。 虽然,AAV在组织和生物体中表现出一定程度的扩散(Le Guiner et al .(2011) 4 CN 111601884 A 说 明 书 2/44 页 “Biodistribution and shedding of AAV vectors”Methods in molecular biology 807,339-359),但它们在大多数治疗方法中通常需要局部施用,特别是用于眼部基因治疗。 对于光感受器或视网膜色素上皮(RPE)细胞的转导,将AAV颗粒施用于视网膜下腔(即,在 RPE和光感受器之间注射液体后形成的空腔)(Mühlfriedel et al .(2013)“Optimized technique for subretinal injections in mice”Methods in molecular biology 935, 343-349,Ochakovski et al.(2017)“Retinal Gene Therapy:Surgical Vector Delivery in the Translation to Clinical Trials.”Front Neurosci 11:174)。 结构上,AAV是小的(25nm)、无包膜病毒,有二十面体衣壳。在其衣壳(cap)蛋白的 组成和结构上不同的天然存在的或工程化的AAV变体(也是AAV血清型)具有不同的嗜性,即 转导不同(视网膜的)细胞类型的能力(Boye et al.(2013)“A comprehensive review of retinal gene therapy”Molecular therapy 21 509-519and Trapani et al .(2014) “Vector platforms for gene therapy of inherited retinopathies”Prog Retin Eye Res)。当与泛活性启动子结合时,这种嗜性决定了基因表达的位点。然而与细胞类型特异性 启动子结合时,位点特异性的程度(即转基因仅在视杆或视锥光感受器中表达)由AAV血清 型的嗜性和启动子的特异性二者共同来决定( et al .(2015)“Retinal gene delivery by adeno-associated virus(AAV)vectors:Strategies and applications” European journal of pharmaceutics and biopharmaceutics)。向眼睛的玻璃体中施用 AAV导致内视网膜内细胞(主要是神经节细胞和Müller胶质细胞)的转导(Trapani et al. (2014)“Vector platforms for gene therapy of inherited retinopathies”Prog Retin Eye Res)。 由于在内界膜(ILM)的物理屏障或在ILM缺乏合适的受体,当玻璃体内施用时,没 有一种天然存在的AAV血清型能够转导光感受器。 成功的基因治疗方法的先决条件是在靶细胞中高效且持久的转基因表达,仅具有 最小的脱靶表达和最低可能的不良作用。尽管可用的AAV载体构成了有价值的基因递送工 具,但是仍然存在对改进的AAV的开发的强烈需求。 为了靶向光感受器,需要将AAV施用于视网膜下腔。这种侵入性很强的外科手术会 导致视网膜从RPE脱离。视网膜脱离通常是暂时性的,视网膜复位后对视网膜功能和形态没 有任何严重或长期的有害影响。然而,在变性的视网膜和/或如果脱离涉及黄斑中心凹区 域,由于视网膜下注射引起的附带损害的风险可能会增加。因此,需要改进的AAV载体,特别 是关于生物利用度、递送途径和靶细胞特异性。
技术实现要素:
因此,仍然需要开发进一步改进的AAV载体。在此,我们描述了从体内选择进化的 新型AAV,其具有将基因转移到视网膜细胞(尤其是光感受器)中的改进的特性。 本发明符合此需要,如本文中所描述的且如在实施例、附图和权利要求书中所描 述的。 本发明涉及一种腺相关病毒(AAV),所述AAV包含在对应于SEQ ID NO:1的第587和 588位的位置之间的至少6-8个氨基酸的插入物;其中插入序列从N-末端到C-末端具有式I: X1A-X1B-X2-X3-X4-X5-X6-X7 (式I) 5 CN 111601884 A 说 明 书 3/44 页 其中X5选自P(Pro)、L(Leu)和V(Val); 其中X7为R(Arg); 其中X1A、X1B、X2、X3、X4、X6独立地是任何氨基酸; 其中X1A和/或X1B独立地任选地不存在或存在, 或者 其中X5选自S(Ser)和T(Thr); 其中X7为S(Ser); 其中X1A、X1B、X2、X3、X4、X6独立地是任何氨基酸; 其中X1A和/或X1B独立地任选地不存在或存在, 或者 其中X5选自A(Ala)和Q(Gln); 其中X7为A(Ala); 其中X1A、X1B、X2、X3、X4、X6独立地是任何氨基酸; 其中X1A和/或X1B独立地任选地不存在或存在, 其中当将AAV静脉内施用于C57-Bl6J小鼠的尾静脉中时,在24小时后,病毒AAV DNA存在于小鼠视网膜细胞核提取物中,并且 其中当将AAV静脉内施用于C57-Bl6J小鼠的尾静脉中以进行视杆细胞分选,以及 施用于RG-eGFP小鼠品系R685933的尾静脉中以进行视锥细胞分选时,施用后24小时,病毒 AAV DNA存在于使用抗CD73包被的磁珠进行的MAC分选的视杆细胞和/或经FACS分选的视锥 细胞中,其中所述视锥细胞表达eGFP,并基于其eGFP表达被进行FACS分选。 此外,本发明涉及用作药物的本发明的AAV。 此外,本发明涉及用于治疗光感受器疾病的本发明的AAV。 本发明还涉及包含本发明AAV的药物组合物。 此外,本发明涉及本发明的AAV用于视网膜细胞核转导的体外用途。 此外,本发明涉及一种用于筛选插入序列的方法,所述方法包括 i)将AAV文库静脉内施用于受试者中,其中每个AAV包含一种插入序列; ii)从视网膜细胞核提取物中分离病毒AAV DNA; iii)将分离的病毒AAV DNA亚克隆到第二AAV文库中; iv)将如步骤iii)中获得的第二AAV文库静脉内施用于受试者中; v)从视杆或视锥光感受器中分离病毒AAV DNA; vi)确定插入序列的序列, 从而获得插入序列。 本发明还涉及可通过本发明的筛选方法获得的肽(插入序列)。 本发明还涉及包含本发明的AAV的试剂盒。 附图说明 附图显示: 图1:来自整个视网膜的肽插入物 可以鉴定出在AAV2 VP1的第587位具有列出的肽插入物的、被赋予了在静脉内递 6 CN 111601884 A 说 明 书 4/44 页 送后24小时内靶向视网膜细胞并将它们的基因组转移到视网膜细胞核中的能力的下列新 型AAV(仅列出具有2个或更多个NGS读段数的变体,具有20个以上NGS读段数的变体用粗体 字母标记)。 图2:来自视杆细胞光感受器的肽插入物 此外,可以鉴定出在AAV2 VP1的第587位具有列出的肽插入物的、被赋予了在静脉 内递送后24小时内靶向视杆细胞光感受器的能力的下列新型AAV(仅列出具有2个或更多个 NGS读段数的肽,具有20个以上NGS读段数的变体用粗体字母标记)。 图3:来自视锥细胞光感受器的肽插入物 最后,可以鉴定出在AAV2 VP1的第587位具有列出的肽插入物的、被赋予了在静脉 内递送后24小时内靶向视锥细胞光感受器的能力的下列新型AAV(仅列出具有2个或更多个 NGS读段数的肽,具有20个以上NGS读段数的变体用粗体字母标记)。 图4:具有RGD、RGS或NGR基序的鉴定的肽插入物。 一些鉴定的AAV在它们的插入序列中包含推定的整合素结合基序,如RGD、RGS或 NGR。这些插入序列总结于图4中。 图5:在一个以上的组织/细胞群体中鉴定的肽插入物。显示了每个实验中NGS读段 的数量。在一个以上的实验中检测到几个AAV。在图5中总结了这些重叠的AAV,所述AAV包括 具有各自NGS读段数的所示插入序列。 图6:代表性的体内共聚焦扫描激光检眼镜检查(cSLO)图像,其显示在单次玻璃体 内注射1μl(含约2×10E9载体基因组)包装有新型AAV2变体(肽插入序列在每个图像顶部给 出)之一的AAV-sc-CMV-eGFP病毒悬液后2周,10周龄C57-BL6J小鼠的眼底中的eGFP自发荧 光。在多种视网膜细胞类型和大面积视网膜中观察到强烈的eGFP自发荧光(灰色)。 图7:代表性的共聚焦扫描显微镜图像,其显示在单次玻璃体内注射1μl(含约2× 10E9载体基因组)包装有新型AAV2(肽插入序列在每个图像栏的顶部给出)之一的AAV-sc- CMV-eGFP病毒悬液后3周,11周龄C57-BL6J小鼠视网膜横截面中的eGFP荧光。对于变体 “NNPTPSR”(参见上图中的概览图像和左下图中的特写图)和“GLSPPTR”(仅在右下图中显示 特写图),在多种视网膜细胞类型和层中以及在视网膜横截面的整个区域中观察到强烈的 eGFP荧光(灰色)。特别地,许多光感受器内节(is)和外核层(onl)内的细胞体是eGFP阳性, inl,内核层;gel,神经节细胞层。比例尺在上图中标记100μm,在下图中标记25μm。 图8:代表性的共聚焦扫描显微镜图像,其显示在单次视网膜下(上图)或玻璃体内 (下图)注射1μl(含约2×10E9载体基因组)包装有新型AAV2(肽插入序列在每个图像栏的顶 部给出)之一的AAV-sc-CMV-eGFP病毒悬液后3周,11周龄C57-BL6J小鼠的视网膜横截面中 的eGFP荧光。在多种视网膜细胞类型和层中观察到强烈的eGFP荧光(灰色)。特别地,许多光 感受器内节(is)和外核层(onl)内的细胞体是eGFP阳性的。视网膜下注射后,在视网膜色素 上皮细胞(rpe)中也观察到强烈的eGFP荧光。inl,内核层;gel,神经节细胞层。比例尺标记 25pm。 图9:(A)代表性的体内共聚焦扫描激光检眼镜检查(cSLO)图像,其显示在单次玻 璃体内注射(WPI)1μl(含约2×10E9载体基因组)包装有新型AAV变体(GLSPPTR或NNPTPSR; 肽插入序列在每个图像顶部给出)之一的AAV-sc-CMV-eGFP病毒悬液后1、2和3周,2月龄 C57-BL6J小鼠的眼底中的eGFP自发荧光。在多种视网膜细胞类型和大面积视网膜中观察到 7 CN 111601884 A 说 明 书 5/44 页 强烈的eGFP自发荧光(灰色)。所有图像都是用相同的激光和探测器设置采集的。(B)代表性 的共聚焦扫描显微镜图像,其显示在单次玻璃体内注射(WPI)1μl(含约2×10E9载体基因 组)包装有AAV2或新型AAV变体(GLSPPTR或NNPTPSR;肽插入序列在每个图像栏的顶部给出) 之一的AAV-sc-CMV-eGFP病毒悬液后3周,在2月龄C57-BL6J小鼠的视网膜横截面中用抗 eGFP抗体获得的免疫信号(灰色)。对于新型变体GLSPPTR和NNPTPSR,分别在多种视网膜细 胞类型和层中观察到强烈的eGFP免疫信号(灰色)。相反,AAV2导致稀疏细胞的较弱的标记。 所有图像都是用相同的激光和探测器设置采集的。特别地,外核层(onl)内的许多光感受器 细胞体是eGFP阳性的。inl,内核层;gcl,神经节细胞层。 图10:代表性的共聚焦扫描显微镜概览图像,其显示在单次玻璃体内注射(WPI)1μ l(含约2×10E9载体基因组)包装有AAV2或新型AAV变体(GLSPPTR或NNPTPSR;肽插入序列在 每个图像栏的顶部给出)之一的AAV-sc-CMV-eGFP病毒悬液后3周,在2月龄C57-BL6J小鼠的 视网膜横截面中用抗eGFP抗体获得的免疫信号(上图,灰色)。对于新型变体GLSPPTR和 NNPTPSR,分别在多种视网膜细胞类型和层中,以及在整个视网膜横截面中观察到强烈的 eGFP免疫信号(灰色)。相反,AAV2导致在视网膜的较小区域中的较弱标记。所有图像都是用 相同的激光和探测器设置采集的。相应的Hoechst 33342核细胞染色(灰色)图像显示在每 张eGFP图像下方。 图11:AAV载体在661w细胞中的转导效率。(A)在用包装有AAV2、7m8或新型AAV变体 (GLSPPTR或NNPTPSR;肽插入序列在每个图像栏的顶部给出)之一的MOI 100,000的AAV-sc- CMV-eGFP转导后48小时,来自661w细胞培养物的代表性落射荧光图像。(B)显示用FACS测量 的、在未转导的661w细胞培养物和用包装有AAV2、7m8或新型AAV变体(GLSPPTR或NNPTPSR; 肽插入序列在每个图像栏的顶部给出)之一的MOI 100,000的AAV-sc-CMV-eGFP转导后48小 时的661w细胞培养物中的相对eGFP强度%的图。 图12:代表性的共聚焦扫描显微镜图像,其显示在单次玻璃体内注射1μl(含约2× 10E9载体基因组)包装有新型AAV 2“GLSPPTR”衣壳的ss-mSWS-eGFP病毒悬液后6周,10周龄 C57-BL6J小鼠视网膜横截面上的eGFP的视锥细胞光感受器表达。AAV-ss-mSWS-eGFP在小鼠 S视蛋白启动子的控制下驱动eGFP的表达。在视锥细胞光感受器中观察到强烈的抗eGFP免 疫信号(灰色)。用视锥细胞标记视锥抑制素(cone arrestin)双重标记的定量显示,78.7± 3.1%(n=4)的视锥抑制素阳性的视锥细胞为eGFP阳性。OS/IS,外节和内节。ONL,外核层。 INL,内核层。OPL,外丛状层。 图13:代表性的共聚焦扫描显微镜图像,其显示在用包装有新型AAV2(肽插入序列 在每个图像栏的顶部给出)之一的sc-CMV-eGFP的1×10E11的总vg(病毒基因组)转导后9 天,在体外(DIV)在人视网膜外植体中AAV介导的eGFP表达。在多种视网膜细胞类型和层中 观察到强烈的天然eGFP荧光(灰色)。特别地,许多光感受器内节(is)和外核层(onl)内的细 胞体是eGFP阳性的。IS,内节。ONL,外核层。INL,内核层。ONL,内核层。GCL,神经节细胞层。 图14:在单次玻璃体内注射1μl(含约1×10E10总vg)包装有新型AAV2“GLSPPTR”衣 壳的ss-mSWS-mCnga3-WPRE(Michalakis et al.(2010)Restoration of Cone Vision in the CNGA3-/-Mouse Model of Congenital Complete Lack of Cone Photoreceptor Function Mol Ther.:2057–2063)后2个月,来自3月龄Cnga3缺陷小鼠的代表性的明视闪烁 ERG测量。显示了在3.1log cd sec/m2和指示频率下处理的眼(黑色)和未处理的对照眼(虚 8 CN 111601884 A 说 明 书 6/44 页 线灰色)的迹线。 图15:代表性的共聚焦扫描显微镜图像,其显示在单次玻璃体内注射1μl(含约1× 10E10总vg)包装有新型AAV2“GLSPPTR”衣壳的ss-mSWS-mCnga3-WPRE(Michalakis et al. (2010)Restoration of Cone Vision in the CNGA3-/-Mouse Model of Congenital Complete Lack of Cone Photoreceptor Function Mol Ther.:2057–2063)后2个月,来自 3月龄Cnga3缺陷小鼠的视网膜横截面上的视锥细胞光感受器表达CNGA3蛋白。在花生凝集 素(PNA)阳性视锥细胞光感受器中发现强烈的抗CNGA3免疫信号。os,外节。onl,外核层。 inl,内核层。gcl,神经节细胞层。 图16:在单次玻璃体内注射1μl(含约1×10E10总vg)的包装有新型AAV2“NNPTPSR” 衣壳的ss-mRho-mCngb1-sv40,(Koch et al.2012)Gene therapy restores vision and delays degeneration in the CNGB1-/-mouse model of retinitis pigmentosa .Hum Mol Genet.:4486–4496)后2个月,来自3月龄Cnga3缺陷小鼠的代表性暗视单闪烁ERG测量。 示出了在指示的光刺激亮度下来自处理的眼(黑色)和未处理的对照眼(虚线灰色)的迹线。
本发明涉及腺相关病毒(AAV),其包括在对应于SEQ ID NO:1的第87和588位的位置之间的至少6‑8个氨基酸的插入物。还预期用作药物的本发明的AAV以及包含本发明的AAV的药物组合物。此外,本发明涉及本发明的AAV用于转导视网膜细胞核的体外用途。还涉及用于筛选插入序列的方 全部
背景技术:
重组腺相关病毒(AAV)载体已被证明是一种非常适合将基因高效且长期转移到视 网膜细胞中的递送系统(Boye et al.(2013)“A comprehensive review of retinal gene therapy”Molecular therapy ,21(2013)509-519and Trapani et al .(2014)“Vector platforms for gene therapy of inherited retinopathies”Prog Retin Eye Res)。AAV 是属于细小病毒科和依赖病毒属的非致病性病毒,仅在存在腺病毒、乳头状瘤病毒或疱疹 病毒的情况下才能复制(Nathwani et al.(2014)“Long-term safety and efficacy of factor IX gene therapy in hemophilia B”N Engl J Med ,371(2014)1994-2004),并具 有大约5kb单链DNA基因组(Trapani et al.(2014)“Vector platforms for gene therapy of inherited retinopathies”Prog Retin Eye Res.)。 AAV的安全性和转基因的长期表达已在大型动物模型、非人灵长类动物和多项人 类试验中被广泛测试(MacLaren et al .(2014)“Retinal gene therapy in patients with choroideremia:initial findings from a phase 1/2clinical trial”Lancet383, 1129-1137 ,Maguire et al .(2008)“Safety and efficacy of gene transfer for Leber 's congenital amaurosis”N Engl J Med ,358 2240-2248 ,Simonelliet al . (2010)”Gene therapy for Leber's congenital amaurosis is safe and effective through1.5years after vector administration”Molecular therapy:the journal of the American Society of Gene Therapy 18,643-650,Nathwani et al.(2014)“Long- term safety and efficacy of factor IX gene therapy in hemophilia B”N Engl J Med 371,1994-2004)。 AAV是用于基因补充疗法(也称为基因增强疗法)的优选基因转移载体。基因补充 疗法是用于治疗遗传疾病的有吸引力的方法,其目的在于恢复由致病突变引起的基因功能 不足或缺乏。目标是恢复受影响的细胞类型的正常生理功能和/或通过补充基因功能来抑 制或延迟变性过程。 基因疗法的目的是恢复由致病突变引起的基因功能不足或缺乏。这种方法对于隐 性遗传疾病的治疗特别有吸引力,在隐性遗传疾病中,异常转录物或蛋白质预计不会干扰 治疗。目标是通过在适当的位置补充足量的正常基因功能来阻止或防止变性过程,并恢复 受影响细胞类型或组织的正常生理功能。 虽然,AAV在组织和生物体中表现出一定程度的扩散(Le Guiner et al .(2011) 4 CN 111601884 A 说 明 书 2/44 页 “Biodistribution and shedding of AAV vectors”Methods in molecular biology 807,339-359),但它们在大多数治疗方法中通常需要局部施用,特别是用于眼部基因治疗。 对于光感受器或视网膜色素上皮(RPE)细胞的转导,将AAV颗粒施用于视网膜下腔(即,在 RPE和光感受器之间注射液体后形成的空腔)(Mühlfriedel et al .(2013)“Optimized technique for subretinal injections in mice”Methods in molecular biology 935, 343-349,Ochakovski et al.(2017)“Retinal Gene Therapy:Surgical Vector Delivery in the Translation to Clinical Trials.”Front Neurosci 11:174)。 结构上,AAV是小的(25nm)、无包膜病毒,有二十面体衣壳。在其衣壳(cap)蛋白的 组成和结构上不同的天然存在的或工程化的AAV变体(也是AAV血清型)具有不同的嗜性,即 转导不同(视网膜的)细胞类型的能力(Boye et al.(2013)“A comprehensive review of retinal gene therapy”Molecular therapy 21 509-519and Trapani et al .(2014) “Vector platforms for gene therapy of inherited retinopathies”Prog Retin Eye Res)。当与泛活性启动子结合时,这种嗜性决定了基因表达的位点。然而与细胞类型特异性 启动子结合时,位点特异性的程度(即转基因仅在视杆或视锥光感受器中表达)由AAV血清 型的嗜性和启动子的特异性二者共同来决定( et al .(2015)“Retinal gene delivery by adeno-associated virus(AAV)vectors:Strategies and applications” European journal of pharmaceutics and biopharmaceutics)。向眼睛的玻璃体中施用 AAV导致内视网膜内细胞(主要是神经节细胞和Müller胶质细胞)的转导(Trapani et al. (2014)“Vector platforms for gene therapy of inherited retinopathies”Prog Retin Eye Res)。 由于在内界膜(ILM)的物理屏障或在ILM缺乏合适的受体,当玻璃体内施用时,没 有一种天然存在的AAV血清型能够转导光感受器。 成功的基因治疗方法的先决条件是在靶细胞中高效且持久的转基因表达,仅具有 最小的脱靶表达和最低可能的不良作用。尽管可用的AAV载体构成了有价值的基因递送工 具,但是仍然存在对改进的AAV的开发的强烈需求。 为了靶向光感受器,需要将AAV施用于视网膜下腔。这种侵入性很强的外科手术会 导致视网膜从RPE脱离。视网膜脱离通常是暂时性的,视网膜复位后对视网膜功能和形态没 有任何严重或长期的有害影响。然而,在变性的视网膜和/或如果脱离涉及黄斑中心凹区 域,由于视网膜下注射引起的附带损害的风险可能会增加。因此,需要改进的AAV载体,特别 是关于生物利用度、递送途径和靶细胞特异性。
技术实现要素:
因此,仍然需要开发进一步改进的AAV载体。在此,我们描述了从体内选择进化的 新型AAV,其具有将基因转移到视网膜细胞(尤其是光感受器)中的改进的特性。 本发明符合此需要,如本文中所描述的且如在实施例、附图和权利要求书中所描 述的。 本发明涉及一种腺相关病毒(AAV),所述AAV包含在对应于SEQ ID NO:1的第587和 588位的位置之间的至少6-8个氨基酸的插入物;其中插入序列从N-末端到C-末端具有式I: X1A-X1B-X2-X3-X4-X5-X6-X7 (式I) 5 CN 111601884 A 说 明 书 3/44 页 其中X5选自P(Pro)、L(Leu)和V(Val); 其中X7为R(Arg); 其中X1A、X1B、X2、X3、X4、X6独立地是任何氨基酸; 其中X1A和/或X1B独立地任选地不存在或存在, 或者 其中X5选自S(Ser)和T(Thr); 其中X7为S(Ser); 其中X1A、X1B、X2、X3、X4、X6独立地是任何氨基酸; 其中X1A和/或X1B独立地任选地不存在或存在, 或者 其中X5选自A(Ala)和Q(Gln); 其中X7为A(Ala); 其中X1A、X1B、X2、X3、X4、X6独立地是任何氨基酸; 其中X1A和/或X1B独立地任选地不存在或存在, 其中当将AAV静脉内施用于C57-Bl6J小鼠的尾静脉中时,在24小时后,病毒AAV DNA存在于小鼠视网膜细胞核提取物中,并且 其中当将AAV静脉内施用于C57-Bl6J小鼠的尾静脉中以进行视杆细胞分选,以及 施用于RG-eGFP小鼠品系R685933的尾静脉中以进行视锥细胞分选时,施用后24小时,病毒 AAV DNA存在于使用抗CD73包被的磁珠进行的MAC分选的视杆细胞和/或经FACS分选的视锥 细胞中,其中所述视锥细胞表达eGFP,并基于其eGFP表达被进行FACS分选。 此外,本发明涉及用作药物的本发明的AAV。 此外,本发明涉及用于治疗光感受器疾病的本发明的AAV。 本发明还涉及包含本发明AAV的药物组合物。 此外,本发明涉及本发明的AAV用于视网膜细胞核转导的体外用途。 此外,本发明涉及一种用于筛选插入序列的方法,所述方法包括 i)将AAV文库静脉内施用于受试者中,其中每个AAV包含一种插入序列; ii)从视网膜细胞核提取物中分离病毒AAV DNA; iii)将分离的病毒AAV DNA亚克隆到第二AAV文库中; iv)将如步骤iii)中获得的第二AAV文库静脉内施用于受试者中; v)从视杆或视锥光感受器中分离病毒AAV DNA; vi)确定插入序列的序列, 从而获得插入序列。 本发明还涉及可通过本发明的筛选方法获得的肽(插入序列)。 本发明还涉及包含本发明的AAV的试剂盒。 附图说明 附图显示: 图1:来自整个视网膜的肽插入物 可以鉴定出在AAV2 VP1的第587位具有列出的肽插入物的、被赋予了在静脉内递 6 CN 111601884 A 说 明 书 4/44 页 送后24小时内靶向视网膜细胞并将它们的基因组转移到视网膜细胞核中的能力的下列新 型AAV(仅列出具有2个或更多个NGS读段数的变体,具有20个以上NGS读段数的变体用粗体 字母标记)。 图2:来自视杆细胞光感受器的肽插入物 此外,可以鉴定出在AAV2 VP1的第587位具有列出的肽插入物的、被赋予了在静脉 内递送后24小时内靶向视杆细胞光感受器的能力的下列新型AAV(仅列出具有2个或更多个 NGS读段数的肽,具有20个以上NGS读段数的变体用粗体字母标记)。 图3:来自视锥细胞光感受器的肽插入物 最后,可以鉴定出在AAV2 VP1的第587位具有列出的肽插入物的、被赋予了在静脉 内递送后24小时内靶向视锥细胞光感受器的能力的下列新型AAV(仅列出具有2个或更多个 NGS读段数的肽,具有20个以上NGS读段数的变体用粗体字母标记)。 图4:具有RGD、RGS或NGR基序的鉴定的肽插入物。 一些鉴定的AAV在它们的插入序列中包含推定的整合素结合基序,如RGD、RGS或 NGR。这些插入序列总结于图4中。 图5:在一个以上的组织/细胞群体中鉴定的肽插入物。显示了每个实验中NGS读段 的数量。在一个以上的实验中检测到几个AAV。在图5中总结了这些重叠的AAV,所述AAV包括 具有各自NGS读段数的所示插入序列。 图6:代表性的体内共聚焦扫描激光检眼镜检查(cSLO)图像,其显示在单次玻璃体 内注射1μl(含约2×10E9载体基因组)包装有新型AAV2变体(肽插入序列在每个图像顶部给 出)之一的AAV-sc-CMV-eGFP病毒悬液后2周,10周龄C57-BL6J小鼠的眼底中的eGFP自发荧 光。在多种视网膜细胞类型和大面积视网膜中观察到强烈的eGFP自发荧光(灰色)。 图7:代表性的共聚焦扫描显微镜图像,其显示在单次玻璃体内注射1μl(含约2× 10E9载体基因组)包装有新型AAV2(肽插入序列在每个图像栏的顶部给出)之一的AAV-sc- CMV-eGFP病毒悬液后3周,11周龄C57-BL6J小鼠视网膜横截面中的eGFP荧光。对于变体 “NNPTPSR”(参见上图中的概览图像和左下图中的特写图)和“GLSPPTR”(仅在右下图中显示 特写图),在多种视网膜细胞类型和层中以及在视网膜横截面的整个区域中观察到强烈的 eGFP荧光(灰色)。特别地,许多光感受器内节(is)和外核层(onl)内的细胞体是eGFP阳性, inl,内核层;gel,神经节细胞层。比例尺在上图中标记100μm,在下图中标记25μm。 图8:代表性的共聚焦扫描显微镜图像,其显示在单次视网膜下(上图)或玻璃体内 (下图)注射1μl(含约2×10E9载体基因组)包装有新型AAV2(肽插入序列在每个图像栏的顶 部给出)之一的AAV-sc-CMV-eGFP病毒悬液后3周,11周龄C57-BL6J小鼠的视网膜横截面中 的eGFP荧光。在多种视网膜细胞类型和层中观察到强烈的eGFP荧光(灰色)。特别地,许多光 感受器内节(is)和外核层(onl)内的细胞体是eGFP阳性的。视网膜下注射后,在视网膜色素 上皮细胞(rpe)中也观察到强烈的eGFP荧光。inl,内核层;gel,神经节细胞层。比例尺标记 25pm。 图9:(A)代表性的体内共聚焦扫描激光检眼镜检查(cSLO)图像,其显示在单次玻 璃体内注射(WPI)1μl(含约2×10E9载体基因组)包装有新型AAV变体(GLSPPTR或NNPTPSR; 肽插入序列在每个图像顶部给出)之一的AAV-sc-CMV-eGFP病毒悬液后1、2和3周,2月龄 C57-BL6J小鼠的眼底中的eGFP自发荧光。在多种视网膜细胞类型和大面积视网膜中观察到 7 CN 111601884 A 说 明 书 5/44 页 强烈的eGFP自发荧光(灰色)。所有图像都是用相同的激光和探测器设置采集的。(B)代表性 的共聚焦扫描显微镜图像,其显示在单次玻璃体内注射(WPI)1μl(含约2×10E9载体基因 组)包装有AAV2或新型AAV变体(GLSPPTR或NNPTPSR;肽插入序列在每个图像栏的顶部给出) 之一的AAV-sc-CMV-eGFP病毒悬液后3周,在2月龄C57-BL6J小鼠的视网膜横截面中用抗 eGFP抗体获得的免疫信号(灰色)。对于新型变体GLSPPTR和NNPTPSR,分别在多种视网膜细 胞类型和层中观察到强烈的eGFP免疫信号(灰色)。相反,AAV2导致稀疏细胞的较弱的标记。 所有图像都是用相同的激光和探测器设置采集的。特别地,外核层(onl)内的许多光感受器 细胞体是eGFP阳性的。inl,内核层;gcl,神经节细胞层。 图10:代表性的共聚焦扫描显微镜概览图像,其显示在单次玻璃体内注射(WPI)1μ l(含约2×10E9载体基因组)包装有AAV2或新型AAV变体(GLSPPTR或NNPTPSR;肽插入序列在 每个图像栏的顶部给出)之一的AAV-sc-CMV-eGFP病毒悬液后3周,在2月龄C57-BL6J小鼠的 视网膜横截面中用抗eGFP抗体获得的免疫信号(上图,灰色)。对于新型变体GLSPPTR和 NNPTPSR,分别在多种视网膜细胞类型和层中,以及在整个视网膜横截面中观察到强烈的 eGFP免疫信号(灰色)。相反,AAV2导致在视网膜的较小区域中的较弱标记。所有图像都是用 相同的激光和探测器设置采集的。相应的Hoechst 33342核细胞染色(灰色)图像显示在每 张eGFP图像下方。 图11:AAV载体在661w细胞中的转导效率。(A)在用包装有AAV2、7m8或新型AAV变体 (GLSPPTR或NNPTPSR;肽插入序列在每个图像栏的顶部给出)之一的MOI 100,000的AAV-sc- CMV-eGFP转导后48小时,来自661w细胞培养物的代表性落射荧光图像。(B)显示用FACS测量 的、在未转导的661w细胞培养物和用包装有AAV2、7m8或新型AAV变体(GLSPPTR或NNPTPSR; 肽插入序列在每个图像栏的顶部给出)之一的MOI 100,000的AAV-sc-CMV-eGFP转导后48小 时的661w细胞培养物中的相对eGFP强度%的图。 图12:代表性的共聚焦扫描显微镜图像,其显示在单次玻璃体内注射1μl(含约2× 10E9载体基因组)包装有新型AAV 2“GLSPPTR”衣壳的ss-mSWS-eGFP病毒悬液后6周,10周龄 C57-BL6J小鼠视网膜横截面上的eGFP的视锥细胞光感受器表达。AAV-ss-mSWS-eGFP在小鼠 S视蛋白启动子的控制下驱动eGFP的表达。在视锥细胞光感受器中观察到强烈的抗eGFP免 疫信号(灰色)。用视锥细胞标记视锥抑制素(cone arrestin)双重标记的定量显示,78.7± 3.1%(n=4)的视锥抑制素阳性的视锥细胞为eGFP阳性。OS/IS,外节和内节。ONL,外核层。 INL,内核层。OPL,外丛状层。 图13:代表性的共聚焦扫描显微镜图像,其显示在用包装有新型AAV2(肽插入序列 在每个图像栏的顶部给出)之一的sc-CMV-eGFP的1×10E11的总vg(病毒基因组)转导后9 天,在体外(DIV)在人视网膜外植体中AAV介导的eGFP表达。在多种视网膜细胞类型和层中 观察到强烈的天然eGFP荧光(灰色)。特别地,许多光感受器内节(is)和外核层(onl)内的细 胞体是eGFP阳性的。IS,内节。ONL,外核层。INL,内核层。ONL,内核层。GCL,神经节细胞层。 图14:在单次玻璃体内注射1μl(含约1×10E10总vg)包装有新型AAV2“GLSPPTR”衣 壳的ss-mSWS-mCnga3-WPRE(Michalakis et al.(2010)Restoration of Cone Vision in the CNGA3-/-Mouse Model of Congenital Complete Lack of Cone Photoreceptor Function Mol Ther.:2057–2063)后2个月,来自3月龄Cnga3缺陷小鼠的代表性的明视闪烁 ERG测量。显示了在3.1log cd sec/m2和指示频率下处理的眼(黑色)和未处理的对照眼(虚 8 CN 111601884 A 说 明 书 6/44 页 线灰色)的迹线。 图15:代表性的共聚焦扫描显微镜图像,其显示在单次玻璃体内注射1μl(含约1× 10E10总vg)包装有新型AAV2“GLSPPTR”衣壳的ss-mSWS-mCnga3-WPRE(Michalakis et al. (2010)Restoration of Cone Vision in the CNGA3-/-Mouse Model of Congenital Complete Lack of Cone Photoreceptor Function Mol Ther.:2057–2063)后2个月,来自 3月龄Cnga3缺陷小鼠的视网膜横截面上的视锥细胞光感受器表达CNGA3蛋白。在花生凝集 素(PNA)阳性视锥细胞光感受器中发现强烈的抗CNGA3免疫信号。os,外节。onl,外核层。 inl,内核层。gcl,神经节细胞层。 图16:在单次玻璃体内注射1μl(含约1×10E10总vg)的包装有新型AAV2“NNPTPSR” 衣壳的ss-mRho-mCngb1-sv40,(Koch et al.2012)Gene therapy restores vision and delays degeneration in the CNGB1-/-mouse model of retinitis pigmentosa .Hum Mol Genet.:4486–4496)后2个月,来自3月龄Cnga3缺陷小鼠的代表性暗视单闪烁ERG测量。 示出了在指示的光刺激亮度下来自处理的眼(黑色)和未处理的对照眼(虚线灰色)的迹线。